s21#
Avtor: Jakob Urh Veler
Datum izdelave: 2024-05-26
Koda seminarja: S21
Vhodni podatek#
S21#
Rezultati analiz#
Identifikacija proteina#
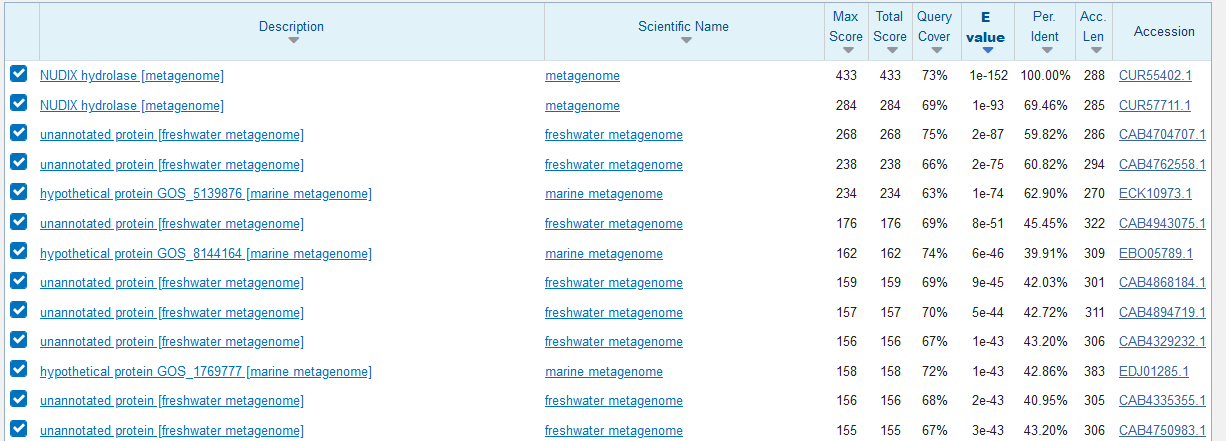
S pomočjo plazmidne karte (https://www.snapgene.com/plasmids/basic_cloning_vectors/pUC57) sem našel multiplo klonirno mesto. Nato sem poravnal multipli klonirni regiji zaporedja (preko Clustal omega) tega plazmida in mojim rekombinantnih plazmidom S21. S tem se je poravnalo le to, kar sem iskal- neporavnan del nukleotidnega zaporedja je zapis za del mojega proteina/vzorca. Nukleotidno zaporedje dela proteina sem vstavil blastx (translirani nukleotidi-proteini) in iskal v zbirki metagenomskih proteinov s21. Protein NUDIX hidrolaza je sestavljen iz 288 aminokislinskih ostankov z zaporedjem: MSNAPTLPDVRAAGAVVTRSGGMVLLVHRPRYDDWSFPKGKLDRGEHVTAAGVREVAEETGLDIRLGPPLRAQRYAAGNRMKTVNYWMGRVIGDDDVSSYVANAEIDEVAWVPFDDAMNLLSYPYDRETLRESTLVRKKTRAFVVLRHAEARARRTWRKNDRLRPLLKPGTLQAQRTVPVLAAYDVTRLASSSSTRCVQTLTPYADVTGYRLRTHDGLSEEDASAASVEKIVSNLVGLNQGVVLCTHRPVLPEVFRALGMSEVRLDPGAMVVAHLRKGKVVATERHGC
{kind=link}
{kind=link}
Določitev strukture in njenih značilnosti#
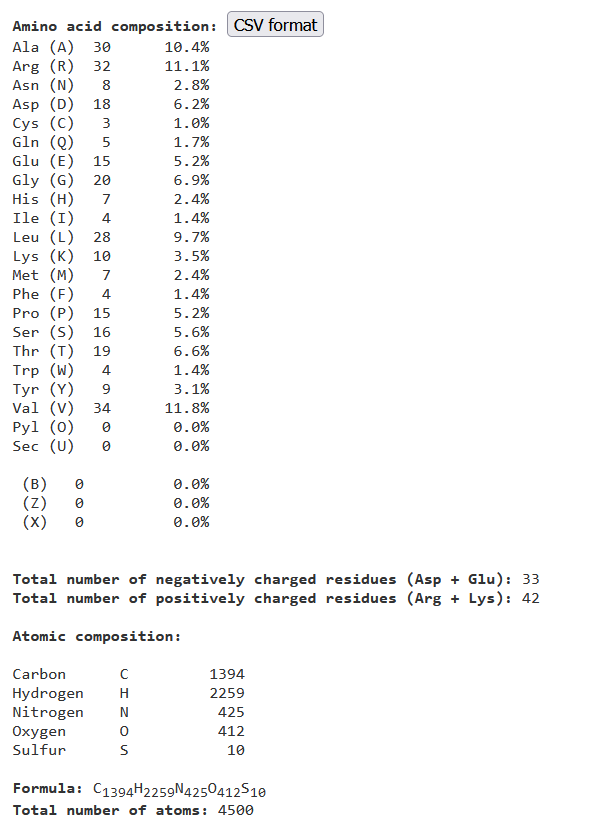
Za analizo aminokislinskih zaporedja vzorca sem uporabil spletno orodje Protparam. Protein vsebuje največ aminokislinskega ostanka valina s21 in več pozitivno nabitih aminokislinskih ostankov. Napovedani ekstincijski koeficient vzorca je 35410L/molcm (reducirani cisteini) s21. Njegova GRAVY vrednost je -0,301, kar pomeni, da je protein hidrofilen (najverjetneje nahaja v vodnem okolju-npr. citoplazma bakterije).
{kind=link}
{kind=link}
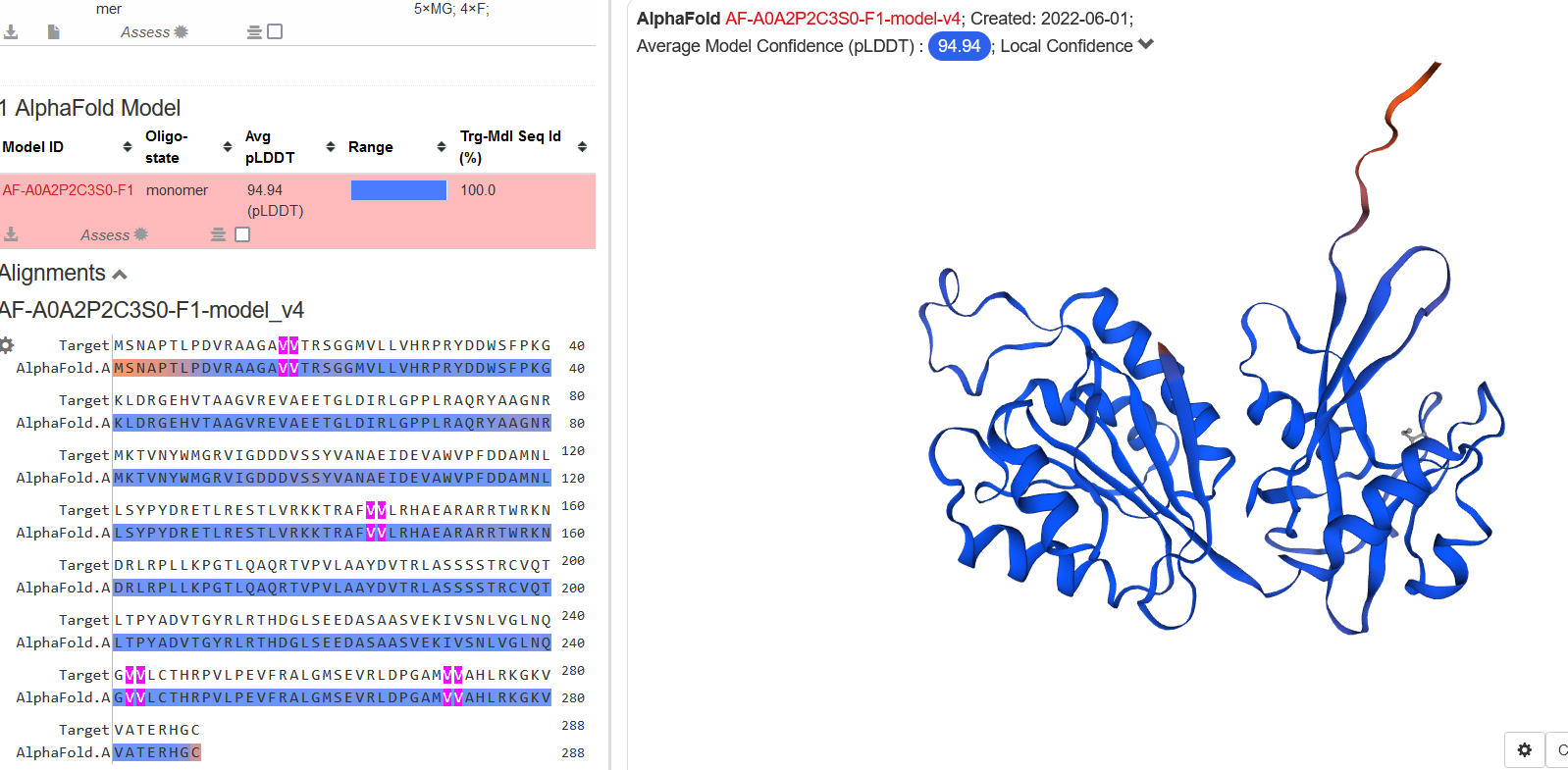
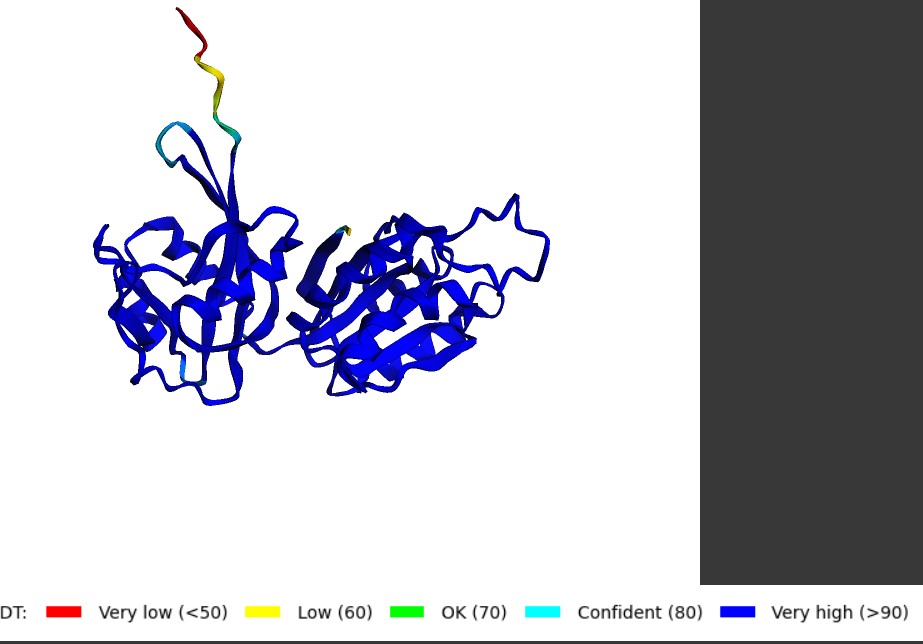
Struktura tega proteina še ni eksperimentalno dokazana (metagenomski izvor), vendar lahko glede na aminokislinsko zaporedje in podobnih struktur v pdb bazi računalniško predvidimo strukturo. Na spletni strani Swiss-model sem vstavil aminokislinsko zaporedje in dobil model s21 in tudi preko biopythona s pomočjo navodil na spletni strani s21. Najmanjša zanesljivost je na N-koncu zaradi visoke gibljivosti tega dela.
{kind=link}
{kind=link}
Da si pomagam s interpretacijo tega modela, sem s pomočjo blastp po nr zbirki poiskal najpodobnejše proteines21. Iz rezultatov lahko z zagotovostjo rečemo, da protein izvira iz bakterije rodu Nocardioides, točne vrste pa ne bi mogel zagotovo določiti. S proteinom iz N. speluncae je 70% identičen v 96% zaporedja. V uniprotu in drugih podatkovnih bazah ni koristnih informacijah o njegovi funkciji, zato sem z blastp po swissprot zbirki poiskal anotirane proteine s21 in naletel na 8-okso-(d)GTP fofatazo (uniprot:A0QUZ2) oziroma v nadaljevanju znan tudi kot anotiran protein, ki izvira iz Mycobacterium smegmatis.
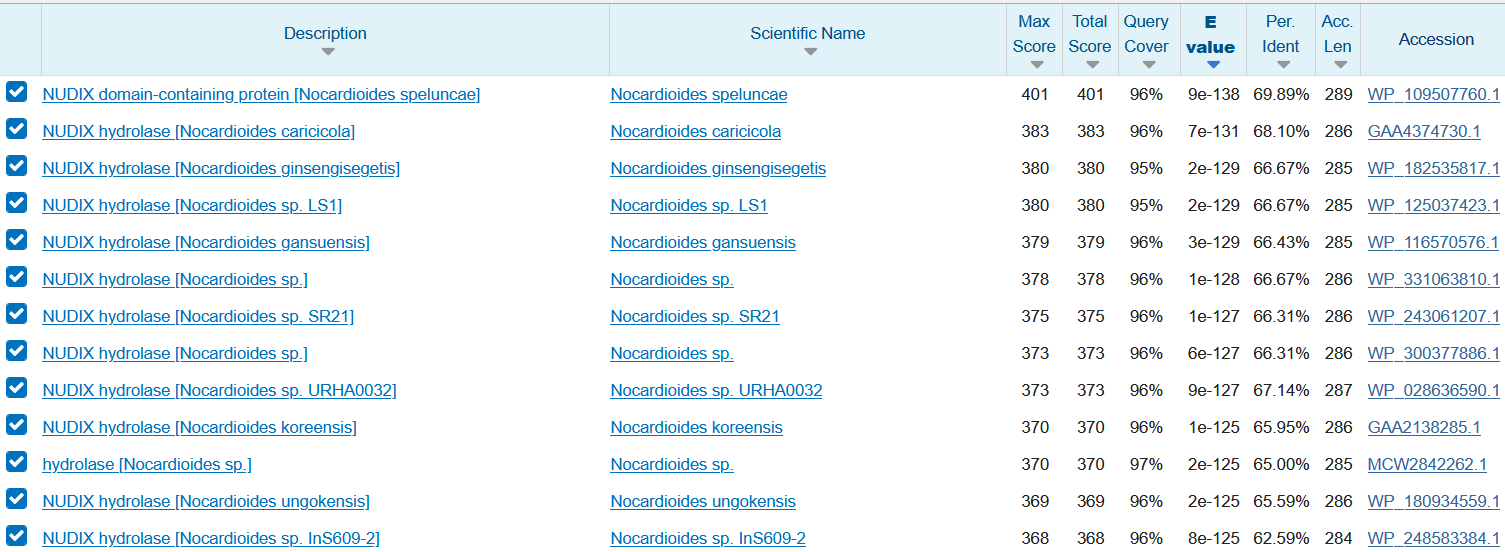{kind=link}
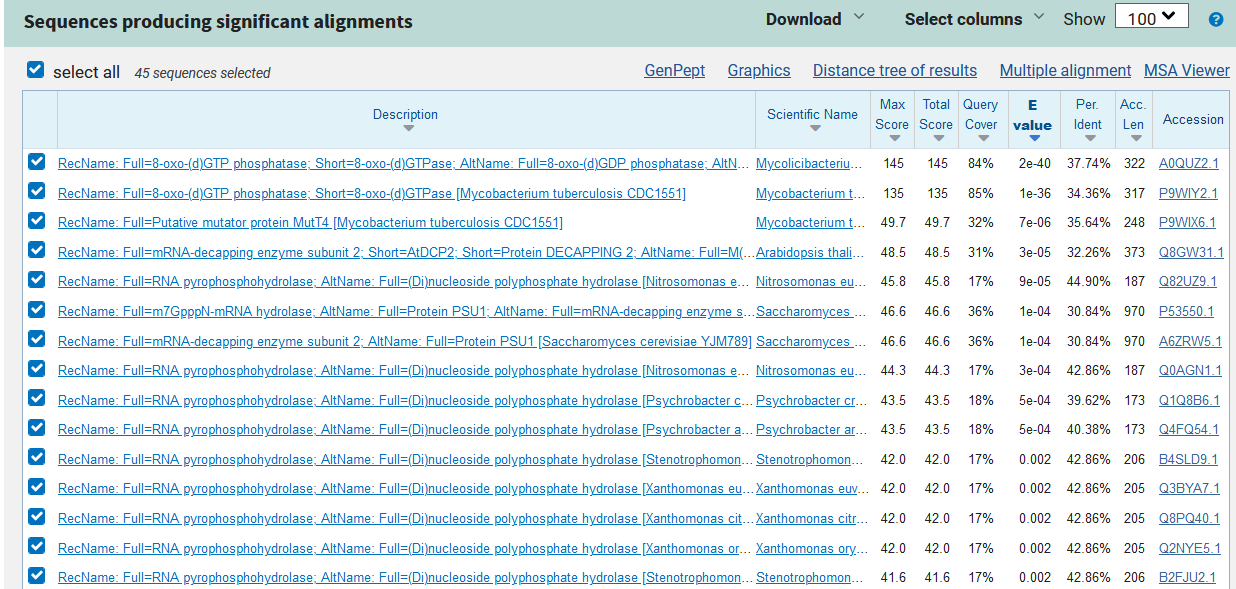{kind=link}
Preden bi lahko naredil povezavo med anotiranim proteinom in mojim vzorcem, sem naredil njuno poravnavo aminokislinskh zaporedij preko omega clustalo s21, poravnavo struktur vzorca (pdb datoteka pridobljena od Swiss-model) in 8-okso-(d)GTP fofataze (pdb:5gg5) v UCSF Chimera [s21](s21-strukturna poravnava_5gg5_v) s21 in primerjavo prisotnost enakih aminokislin v aktivnem mestu s21 s21s21. Obarval sem zeleno aminokislinske ostanke, na katere se veže substrat, in rdeče, ki nase vežejo Mg2+/Mn2+. Roza obarvani ostanki se neujemajo in v oklapaju je zapisan ostanek od anotiranega proteina. Prisotnost pravih aminokislin sem kar ročno preveril. Najprej sem poiskal zaporedje RPRY v vzorcu in od tam naprej označil aminokisline, ki so oddaljene za točno določeno število v anotiranem proteinu. Izkaže se, da je visoka ohranjenost aminokislin v aktivnem mestu in sekundarnih strukturah med vzorcem in 8-okso-(d)GTP fofatazo, zato sem sklepal, da imata enake funkcije, interakcije, lokacijo v celici in posttranslacijske modifikacije. Anotiran protein lahko tvori homodimere, kar bi bilo dobro preveriti tudi pri vzorcu. Če bi želeli to preveriti, bi morali sintetizirati protein in ga izpostaviti raznim eksperimentom-npr. preveriti aktivnost homodimera in monomera.
{kind=link}
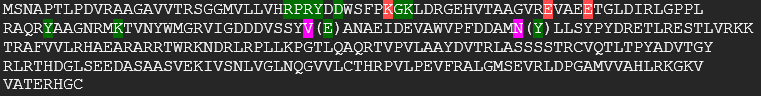{kind=link}
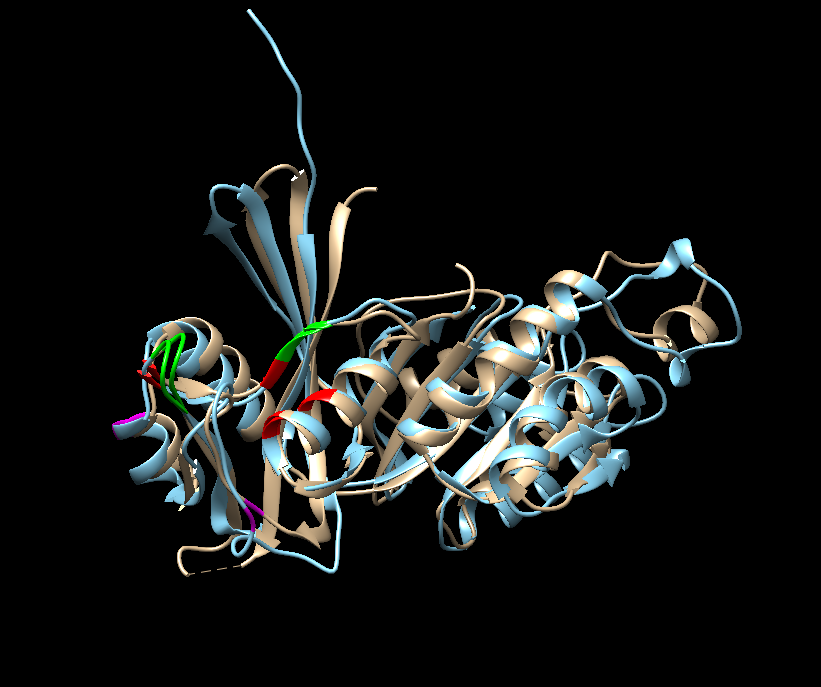{kind=link}
Funkcije proteina#
Torej NUDIX hidrolaza se nahaja v citoplazmi celice, kjer katalizira hidrolizo 8-okso-(d)GTP v 8-okso-(d)GDP in ob visokih koncetracijah encima tudi hidrolizo 8-okso-(d)GDP v 8-okso-(d)GDP. Prav tako je odgovoren za hidrolizo diadenozin polifosfatov ( Ap6A, Ap5A in Ap4A). Encim deluje le ob prisotnosti Mg2+ ali Mn2+. Kataliza je najverjetneje tudi inhibirana s fluoridnimi ioni. Nima posttranslacijskih modifikacij. Zgrajen je iz NUDIX hidrolazne domene (z dobro ohranjenim motivom NUDIX škatle) in neurejeno regijo.
Ohranjenost regij#
S pomočjo clustal omega sem poravnal različne proteine znotraj rodu Nocardioides. Le-te so bili poiskani preko podobnosti z vzorcem blastp znotraj nr zbirke s21s21. Poravnana zaporedja (clustal omega)s21 sem vstavil v Web logo 3 (https://weblogo.threeplusone.com/create.cgi), ki z velikostjo črk aminokislin prikaže, ohranjenost le-tega aminokislinskega ostankas21. Večja kot je črka večja je ohranjenost. Iz rezultatov lahko razberemo: -najbolj ohranjen del zaporedja je VLLVHRPKYDDWSFPKGK, ki je del NUDIX hidrolazne domene, -dobro ohrenjen motiv NUDIX škatle (AAAVREVAEETGL) -aktivna mesta so dobro ohranjena (R<->L pri RPRY vezavnem mestu, zadnja dva aminokislinska ostanka se popolnoma razlikujejo)
{kind=link}
{kind=link}
S pomočjo clustal omega sem poravnal različne proteine med različnimi organizmi (prokarionti in evkarionti). Le-te so bili poiskani preko podobnosti NUDIX domene vzorca blastp znotraj landmark zbirke. Upošteval sem le tiste proteine s21, ki so imeli vrednost E manjšo od 0,0001, ker je vsaka višja vrednost najverjetneje le naključje in nima veze z homolognostjo. Poravnana zaporedja s21 sem vstavil v Web logo 3, ki z velikostjo črk aminokislin prikaže, ohranjenost le-tega ostankas21. Tudi tu sem poravnal celotna zaporedja proteinov, da se vidi, kako je le določen del zaporedja ohranjen. Večja kot je črka večja je ohranjenost. Iz rezultatov lahko razberemo: -da je najbolj ohranjena NUDIX hidrolazna domena skupaj z motivom NUDIX škatle(REV-X-EETG) -domeni N in C konca se zelo razlikujeta med različnimi organizmi
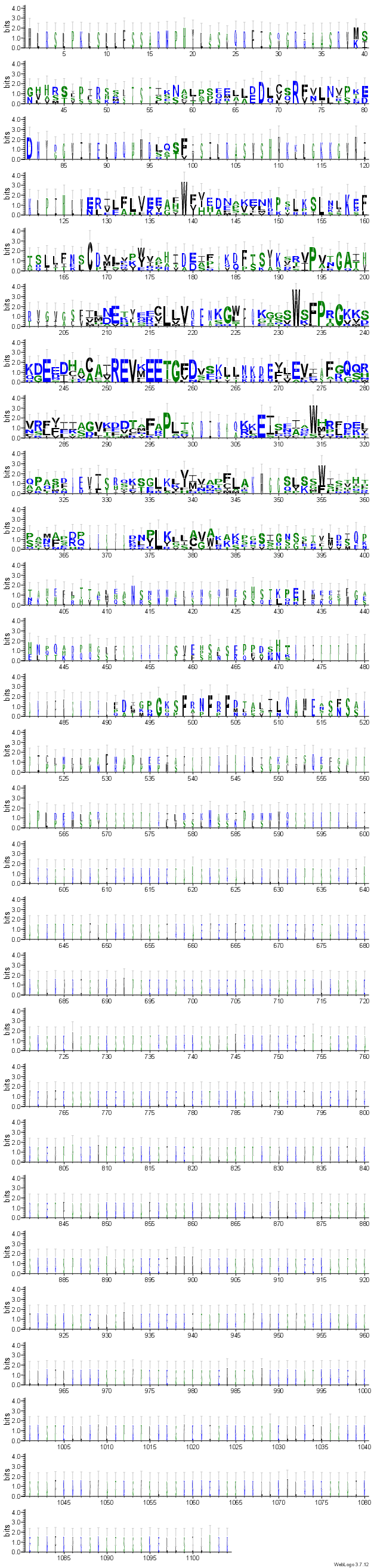{kind=link}
Filogenetsko drevo#
Naredil sem poravnavo tako prokarionskih in evkarionskih podobnih proteinov v clustal omega s21. Dobil sem sledeče filogenetsko drevo s21. Vzorec je najbolj podoben 10N.sp3 in 15N.sp4, kjer je N. okrajšava za rod Nocardioides. Najbližji evkariontski sorodnik je nekaratkeriziran protein iz vinske mušice (Drosophila melanogaster) zaradi bližine v filogenetskem drevesu. Iz Genbanka ne izvem veliko informacij o njegovi funkciji, zato sem s pomočjo blastp po zbirki swissprot poiskal podobne proteine. Izkaže se, da je nekarakteriziran protein najbolj podoben protein 8-okso-dGTP fosfataza NUDT18 (Uniprot: Q568Q0), ki izvira iz navadne cebrice (Danio rerio). Je najbolj podoben evkariontski protein. Katalizira hidroliziro oksidiranih nucleozid difosfatnih derivatov, med katere spadata tudi 8-okso-(d)GTP.
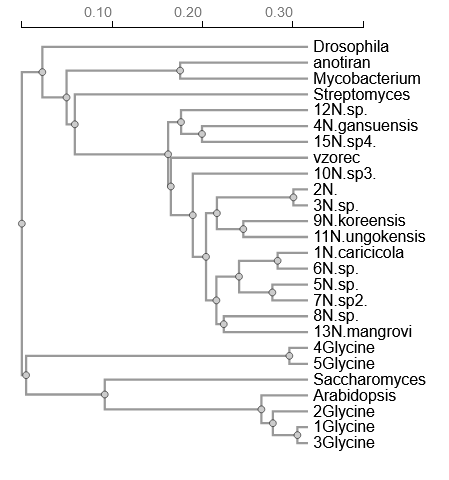{kind=link}
Za analizo interakcij sem uporabil: -prokarionski anotiran protein 8-okso-(d)GTP fofatazo (Uniprot: A0QUZ2), ker je v Swiss-Protu najbolj podoben vzorcu in vsebuje podatke o interakcijah med proteini. Na uniprotu v odseku interakcije je link do spletne strani STRING, ki grafično prikazuje, kateri proteini vplivajo na 8-okso-(d)GTP fofatazo s21. Anotiran protein (rdeče barv, ABK74793.1) interagira z RNazami, DNA polimerazami, fenilalanil-tRNA sintetazami,polifosfat kinazami… -evkarionski anotiran protein 8-okso-dGTP fosfataza NUDT18 (Uniprot: Q568Q0). Sledi podoben postopek kot pri prokarionskem proteinus21. Anotiran protein (rdeče barve, nudt18) interagira z drugimi NUDIX hidrolazami.
{kind=link}
{kind=link}