S10#
Avtor: Dan Kolnik
Datum izdelave: 2024-05-27
Koda seminarja: S10
Vhodni podatek#
Povezava do datoteke z vhodnim podatkom: S10
Rezultati analiz#
Protein patatin#
Iz nukleotidnega zaporedja plazmidnega konstrukta (naslovljenega S10), osnovanega na klonirnem vektorju pUC57, sem identificiral zapis za mikrobni protein ali fragment tega v multipli klonirni regiji. To sem storil tako, da sem na spletu poiskal zaporedje klonirnega vektorja pUC57, potem pa sem zaporedje plazmidnega konstrukta in vektorja pUC57 primerjal med seboj, z ročnim iskanjem. Tako sem ugotovil, kateri del plazmidnega konstrukta predstavlja multipla klonirna regija in kateri del nukleotidnega zaporedja predstavlja zapis za mikrobni protein ali fragment proteina. Zatem sem uporabil orodje blastx, poiskal pa sem metagenomske proteine (env_nr) z nukleotidnim zaporedjem, ki sem ga identificiral v MCS (ang. multiple cloning site) mestu plazmidnega konstrukta. Poiskal sem metagenomske proteine, ker se raziskovalna skupina, katere del sem postal, ukvarja z metagenomiko. Zapis za nukleotidno zaporedje, ki sem ga ustavil za iskanje v orodju, in ga je blastx prevedel v aminokislinsko zaporedje, s katerim je iskal proteine, se je 100% ujemalo (100% identičnost) s proteinom patatin ! blastx, ki je bil določen iz metagenomskega vzorca (mešanica genomov mikroorganizmov iz okolja). Od sedaj naprej bom za protein patatin, ki je bil določen iz metagenomskega vzorca, uporabljal naziv protein patatin. Aminokislinsko zaporedje proteina patatin šteje 273 aminokislinskih ostankov.
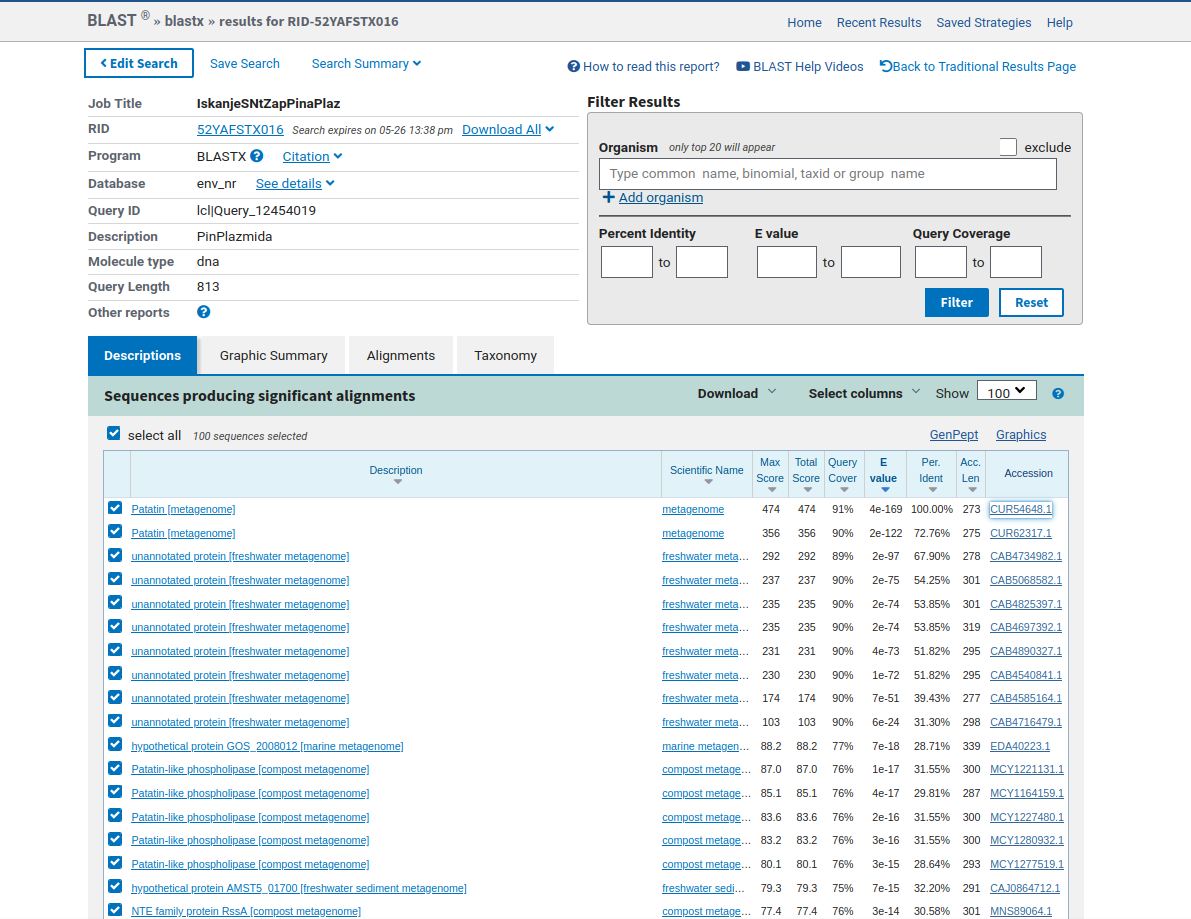{kind=link}
Prokariontski proteini#
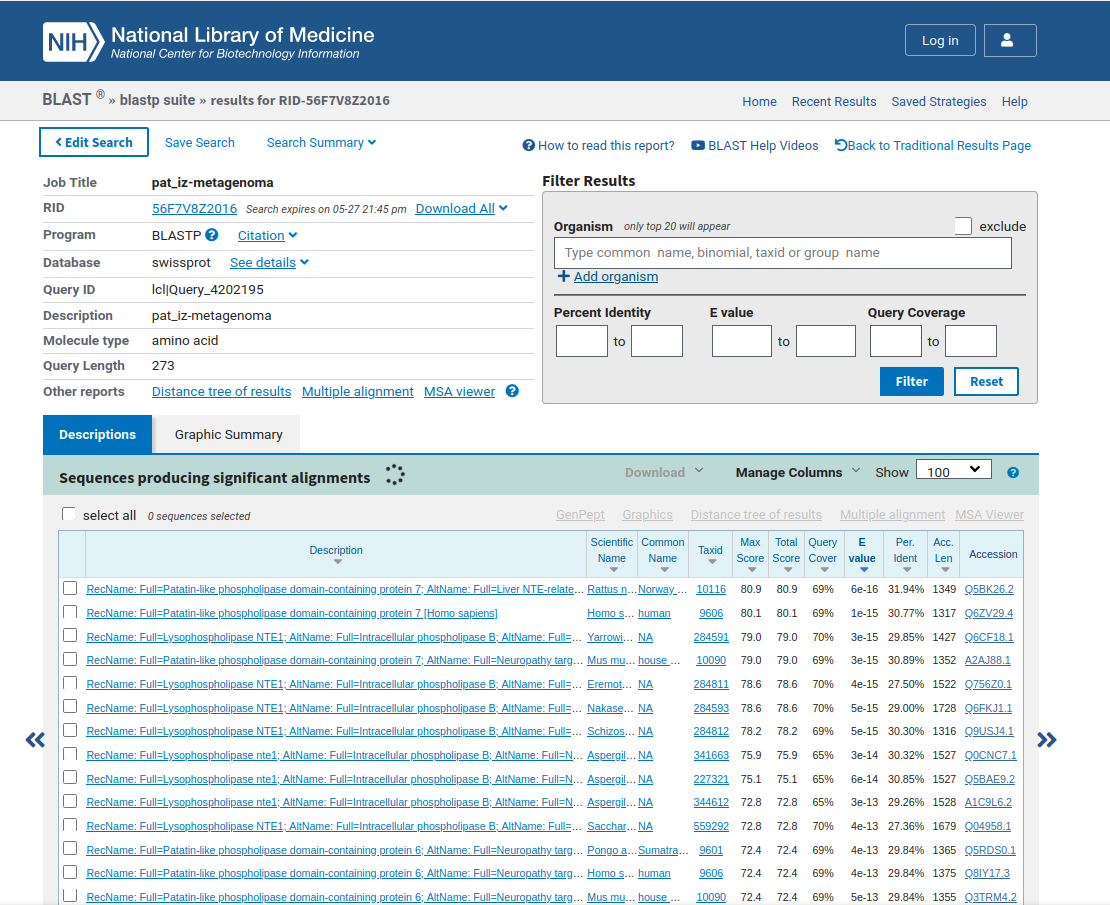
Ker nisem vedel, kako bi o proteinu patatinu več izvedel, sem se odločil, da bom poiskal njemu najbolj podobne proteine. Zato sem z orodjem blastp (ang. protein blast) preko aminokislinskega zaporedja za patatin poiskal “non-redundant protein sequences (nr)” (s tem načinom iskanja sem izločil iskanje “tistega, kar ni potrebno”) ! blastp. Prvi zadetek iskanja je imel najmanjšo E-vrednost. Manjša kot je E-vrednost, bolj podobno je zaporedje. Zaradi tega sem sklepal, da je proteinu patatinu najpodobnejše zaporedje proteina, imenovanega patatinu podobna fosfolipaza (ang. patatin-like phospholipase) iz organizma Actinomycetota bacterium. Na strani podatkovne baze GenBank, ki sem jo odprl iz programa blastp, je pisalo, da je aminokislinsko zaporedje proteina patatinu podobne fosfolipaze iz bakterije Actinomycetota bacterium dolgo 274 aminokislinkih ostankov. Torej, glede na to, da je protein patatin najbolj podoben omenjenu proteinu iz bakterije, se pravi prokariontskega organizma, sem sklepal, da je izvorni organizem proteina patatina bakterija, prokarionstki organizem, natančneje, bakterija Actinomycetota bacterium. Prav tako pa so bili vsi zadetki iskanja z orodjem blastp proteini iz bakterij, in sicer vse te bakterije pripadajo deblu Actinomycetota. Actinomycetota (ali Actinobacteria) je deblo bakterij z zelo raznolikimi člani, ki pa so klasificirani kot Gram-pozitivne bakterije z visokim deležem parov GC. Bakterije tega rodu pripomorejo k razgradnji organskih snovi odmrlih organizmov.
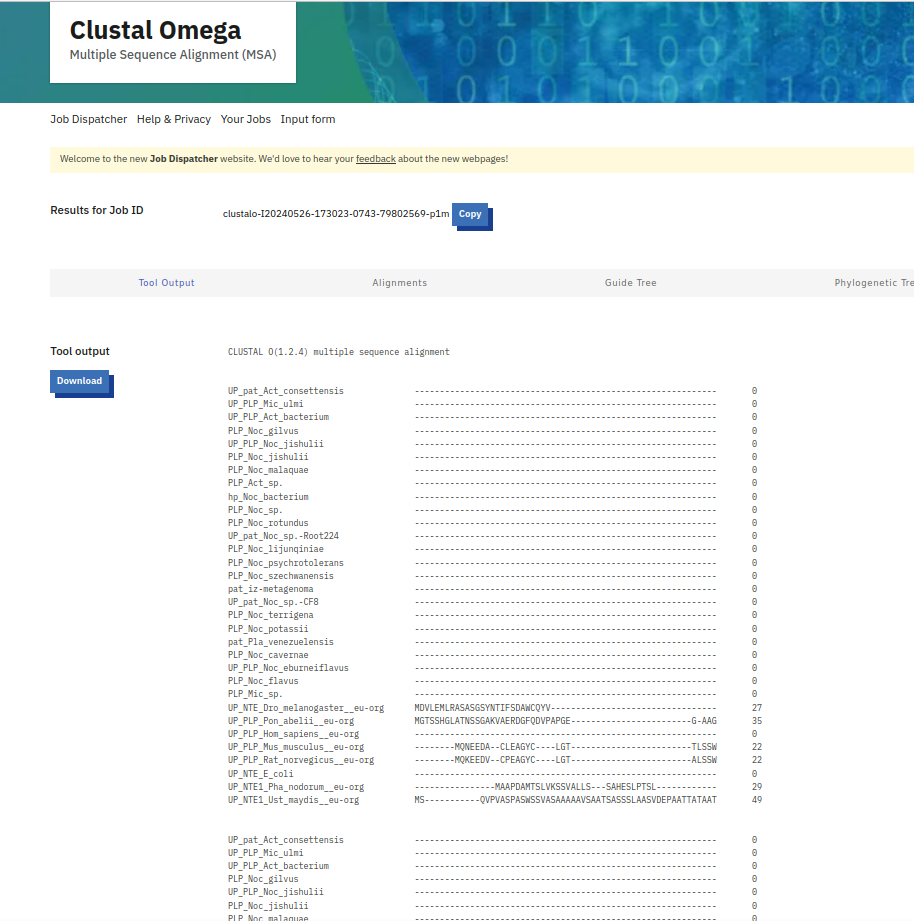
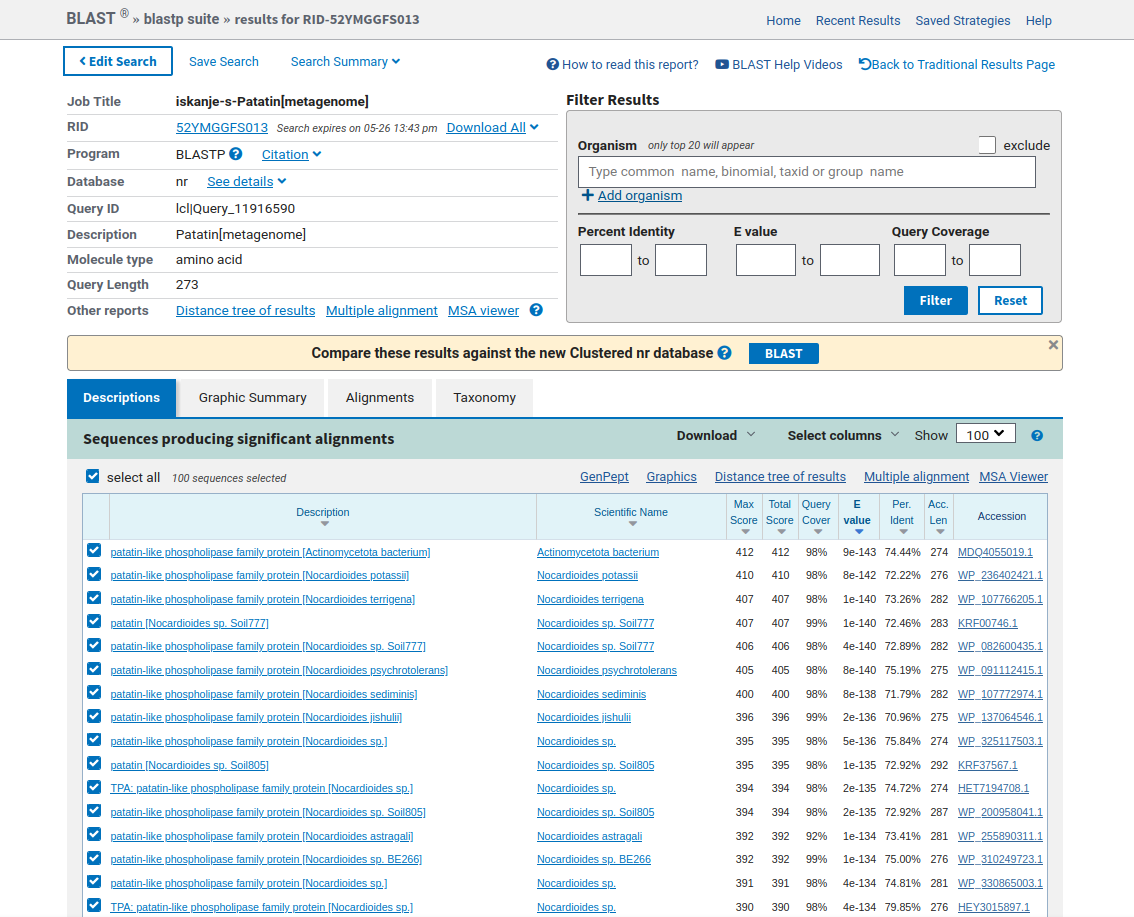{kind=link}
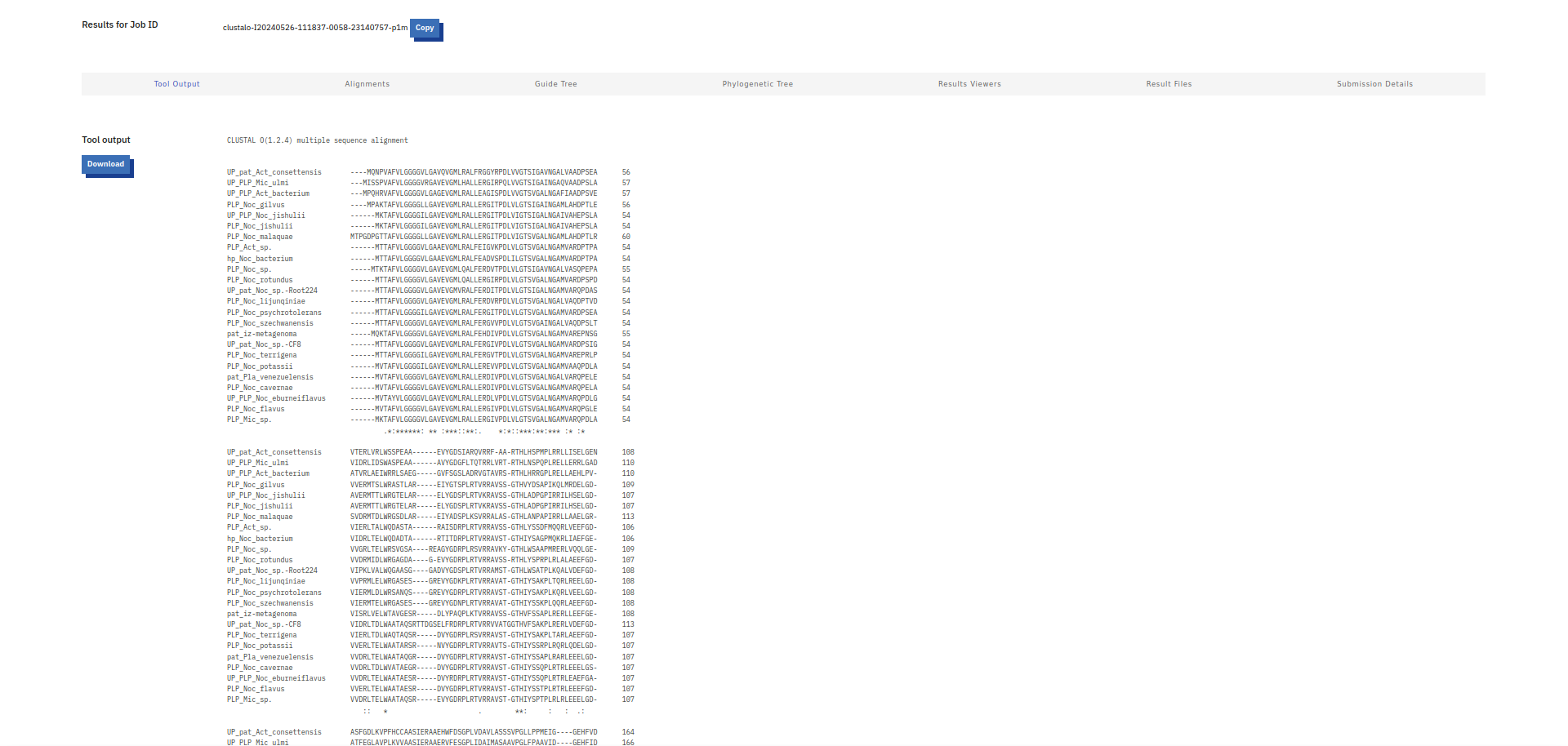
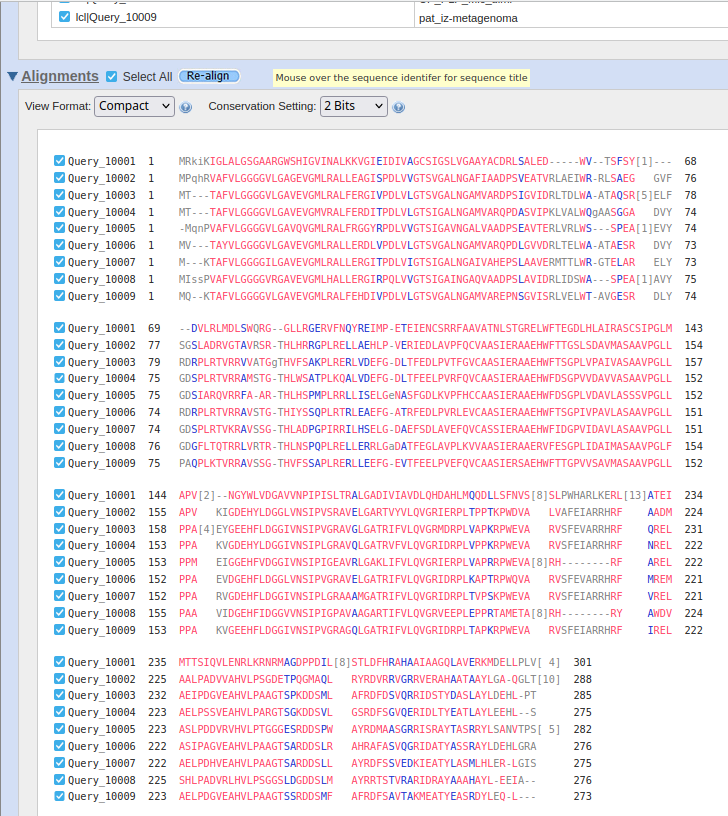
Da bi več izvedel o patatinu najbolj podobnem proteinu, sem na UniProt-u poiskal patatinu podobne proteine iz bakterije Actinomycetota bacterium ! UniProt. Kot rezultat sem dobil 118 različic proteina patatinu podobne fosfolipaze, in želel sem identificirati tisto varianto proteina, ki je patatinu najbolj podobna, zato da bi lahko čim bolje sklepal na lastnosti proteina patatina. Naredil sem več poravnav večih zaporedij (z zaporedji različic proteinov iz bakterije Actinomycetota b. in zaporedjem proteina patatina) z orodjem Clustal Omega. Začel sem s poravnavo 118 različic proteinov skupaj s proteinom patatinom, potem pa sem vsakič izbrisal čim več zaporedij proteinov, ki se niso tako dobro ujemala glede na zaporedje proteina patatina. Na koncu so ostala zaporedja štirih variant proteinov patatinu podobnih fosfolipaz iz bakterije Actinomycetota b., ki so se mi zdela najbolj podobna patatinu ! poravnava. S štirimi najbolj podobni različicami proteina patatinu podobne fosfolipaze sem naredil poravnave, vsako zaporedje posebej sem primerjal s proteinom patatinom z algoritmom globalne poravnave, ki ga uporablja orodje embossNeedle. Ugotovil sem, da je proteinu patatinu, izmed vseh različic proteinov patatinu podobnih fosfolipaz iz bakterije Actinomycetota b. iz UniProt-a, najbolj podobna varianta z 288 aminokislinskimi ostanki (zaporedje ima v delu kode za UniProt znake TK25) ! poravnava_288aa.
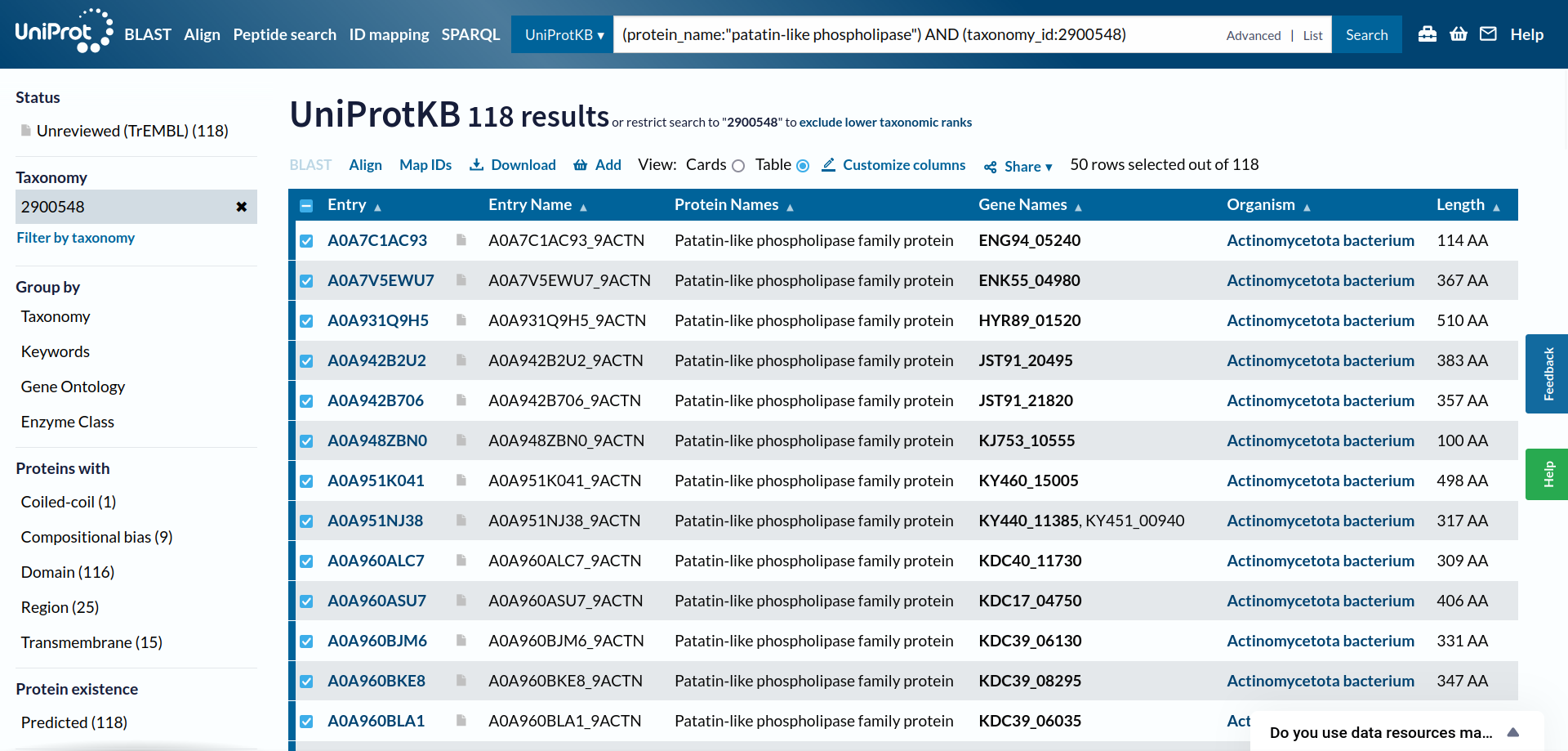{kind=link}
{kind=link}
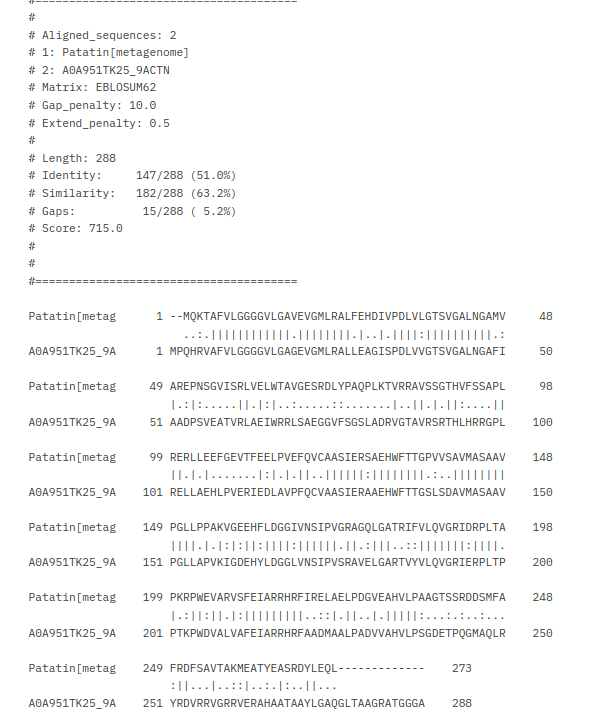{kind=link}
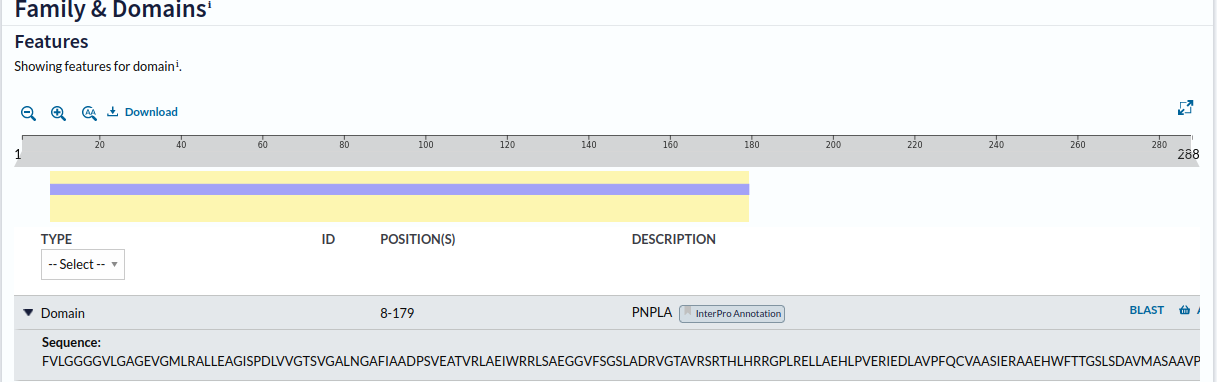
Nato sem za vse štiri zelo podobne različice proteinov na UniProtu preveril, kakšne so njihove lastnosti. Ni bilo veliko podatkov, a očitno tej proteini predstavljajo encime, ker je zapisano, da so sposobni hidrolizirati vezi. Prav tako so vse različice proteina patatinu podobne fosfolipaze udeležene v metabolizmu lipidov. Vse štiri različice vsebujejo samo en poznan motiv sekundarne strukture, in sicer domeno PNPLA, katere regija se začne na oziroma pri N-koncu proteina, predstavlja pa več kot polovico proteina ! domena_PNPLA_pri_288aa. Štiri variante proteinov so med seboj precej podobne. Z oznako PNPLA pa se ne označuje zgolj domene, temveč tudi encime kot take. Iz znanstvenih člankov sem prebral, da so patatinu podobne fosfolipaze (patatin-like phospholipases (PNPLAs)) dobro ohranjeni encimi, ki se nahajajo v prokariontskih in evkariontskih organizmih. Imajo ključno vlogo pri metabolizmu lipidov. Patatinu podobne fosfolipaze katalizirajo hidrolizo sn-2 esterske vezi fosfolipidov, s tem pa lahko povzročijo premik fodfolipidov iz ene strani membrane na drugo, torej lahko spreminjajo sestavo membranskih slojev, lahko pa tudi inducirajo aktivacijo signalov, ki vodijo do programirane celične smrti. Patatinu podobne fosfolipaze interagirajo z membranskimi lipidi, ampak ni nujno, da so del membrane. Iz povedanega do sedaj lahko sklepam, da je torej protein patatin fosfolipaza s podobnimi lastnostmi, kot jih imajo proteini imenovani patatinu podobna fosfolipaza.
{kind=link}
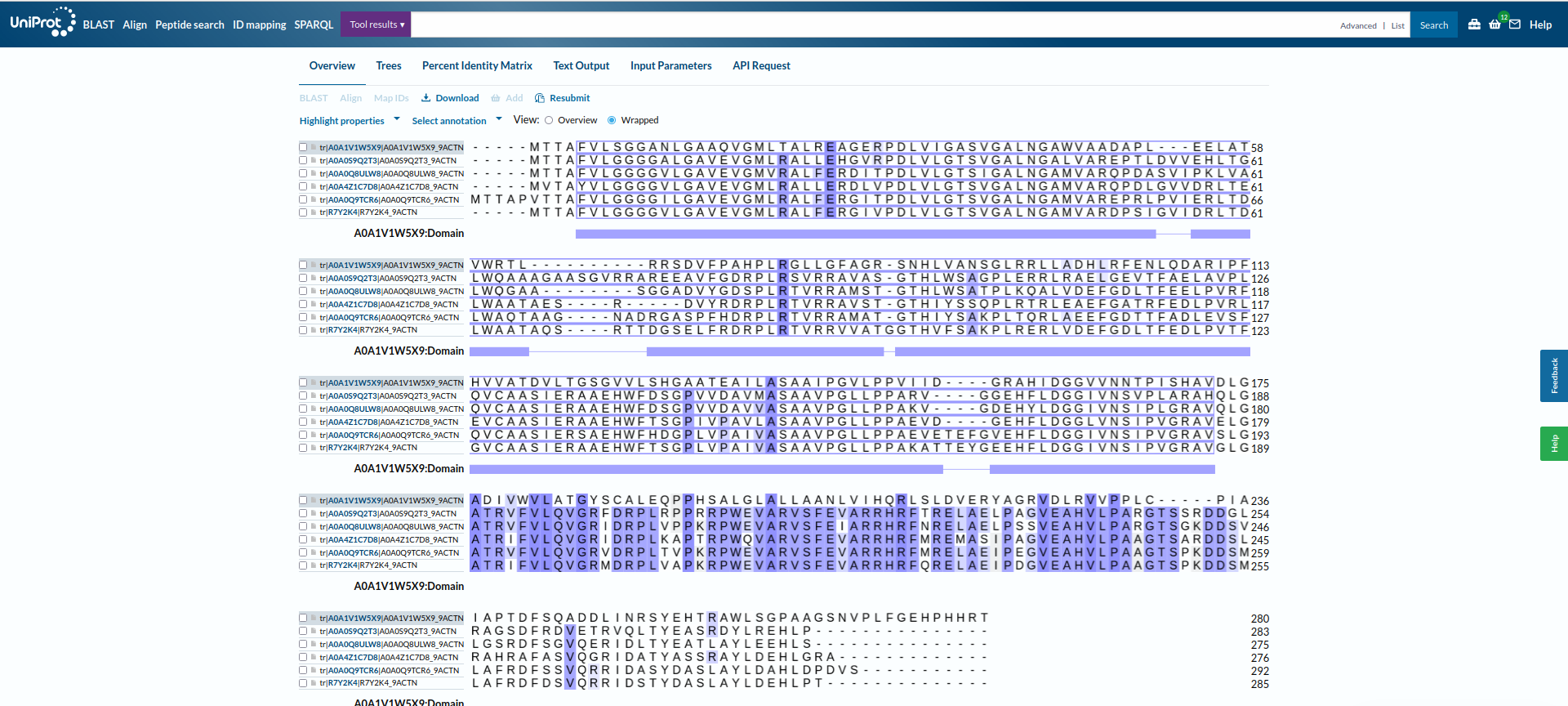
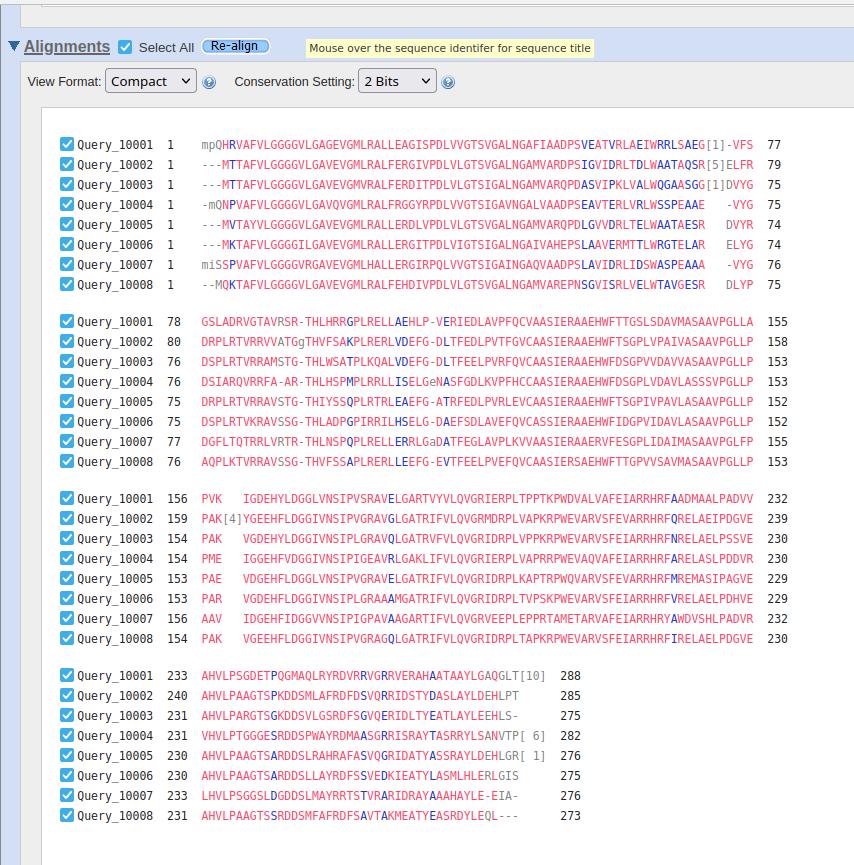
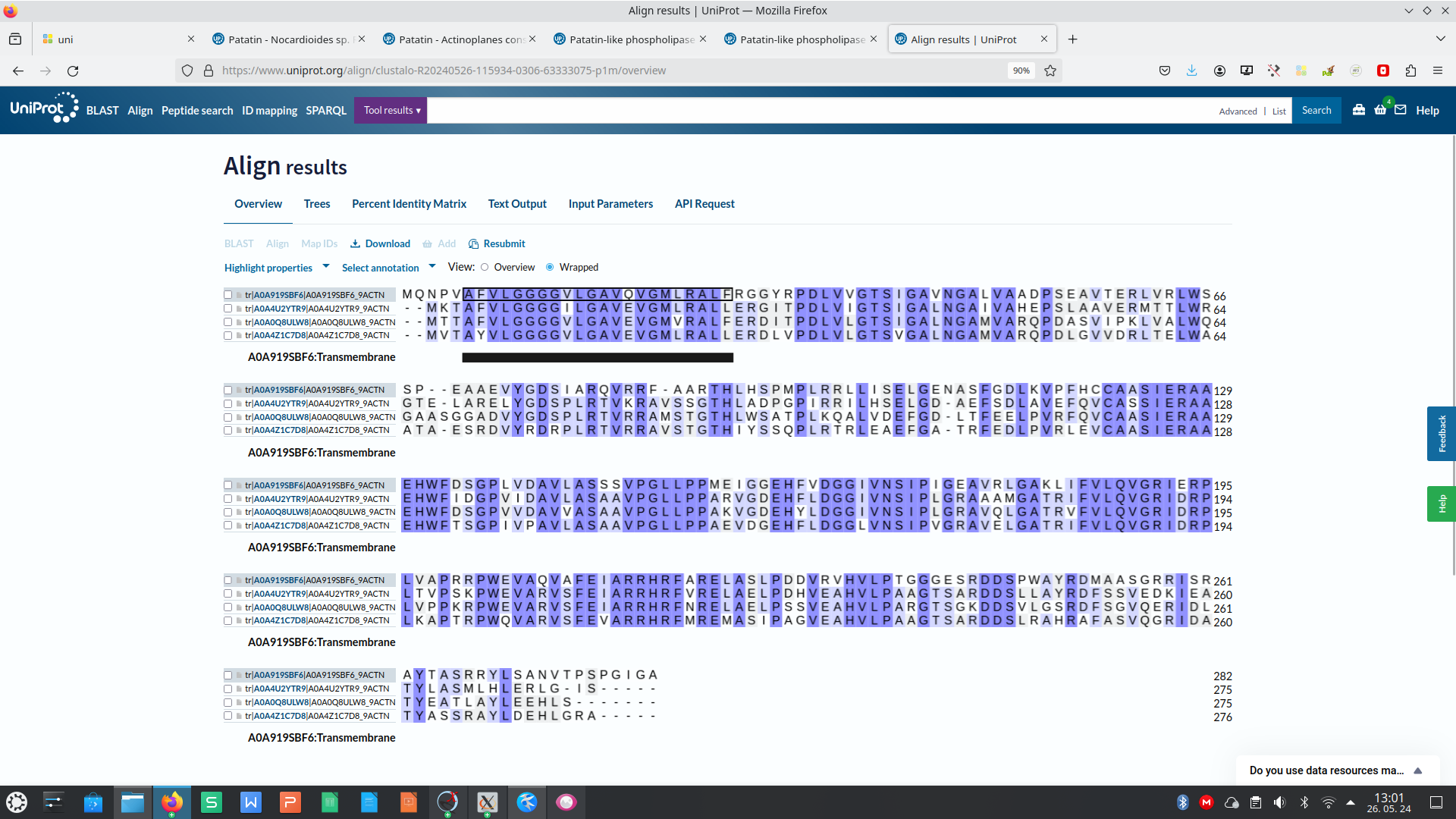
Potem sem na UniProt-u iskal patatinu podobne fosfolipaze oziroma patatinu podobne proteine iz bakterij, katerih proteini so bili najdeni z blastp kot zelo podobni proteini proteinu patatinu. To sem naredil zato, ker sem mislil, da je to najboljša pot, da najdem še več podatkov, da bi lahko primerjal lasnosti in še kaj sklepal za protein patatin. Našel sem nekaj zelo podobnih proteinov iz bakterij iz rodov Nocardioides, Actinoplanes, Plamonospora in Microbacterium. Podobnost vseh proteinov s proteinom patatinom sem preveril z orodjem embossNeedle, ki uporablja algoritem za globalno poravnavo zaporedij.12 proteinov Nocardioides sem tudi poravnaval v UniProt-u s proteinom iz te bakterije, za katerega sem že prej vedel, da je zelo podoben patatinu, potem pa sem glede na poravnavo izbral še dve podobni zaporedji ! poravnava_na_UniProt.
{kind=link}
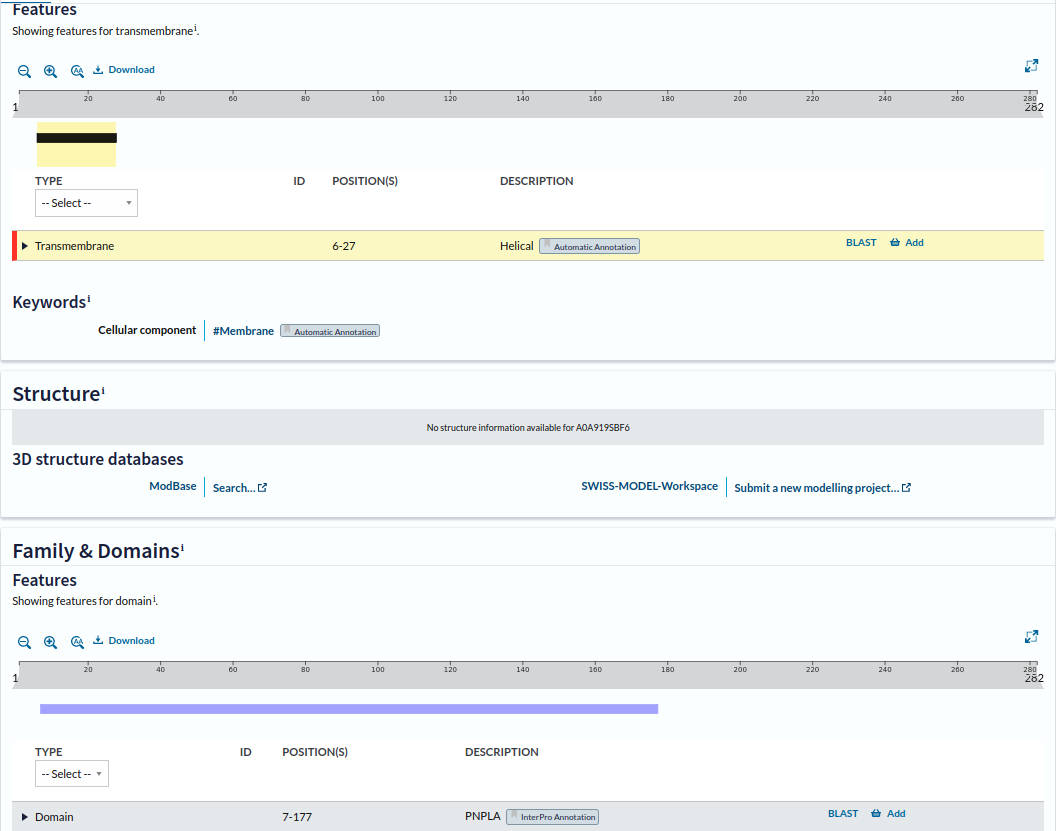
Brez izjeme so vsi patatinu podobni proteini, ki sem jih našel, vsebovali domeno PNPLA. Ta se je nahajala na oziroma pri N-koncu proteinov. Pri zapisu informacij na UniProt-u za bakterijo Actinoplanes consettensis, je pisalo tudi, da se protein patatin v tej bakteriji nahaja v membrani, in sicer od 6. do 27. aminokislinskega ostanka (AFVLGGGGVLGAVQVGMLRALF) se razteza transmembranska regija, ki je v obliki alpha-vijačnice ! transmemb_regija.
{kind=link}
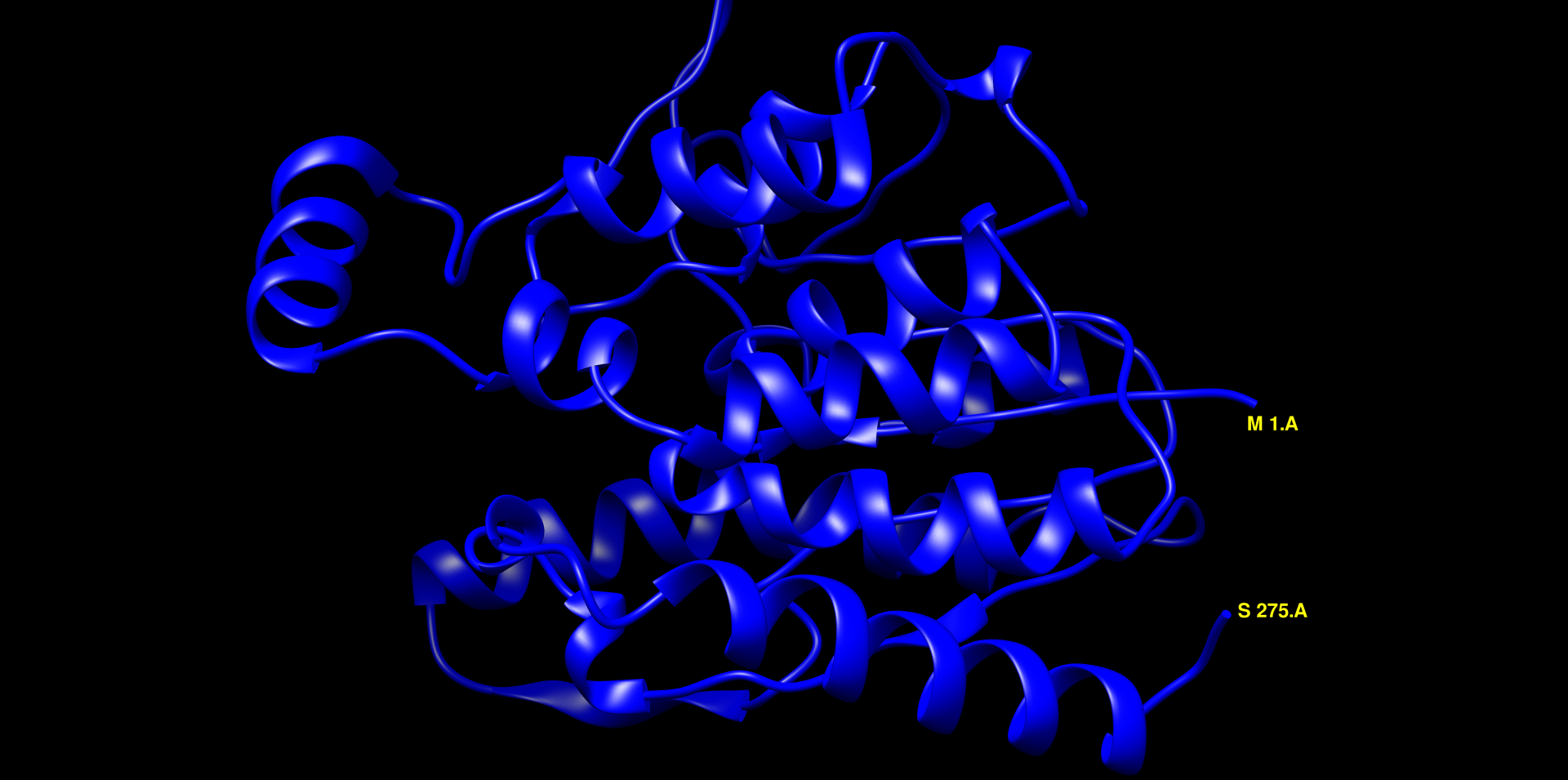
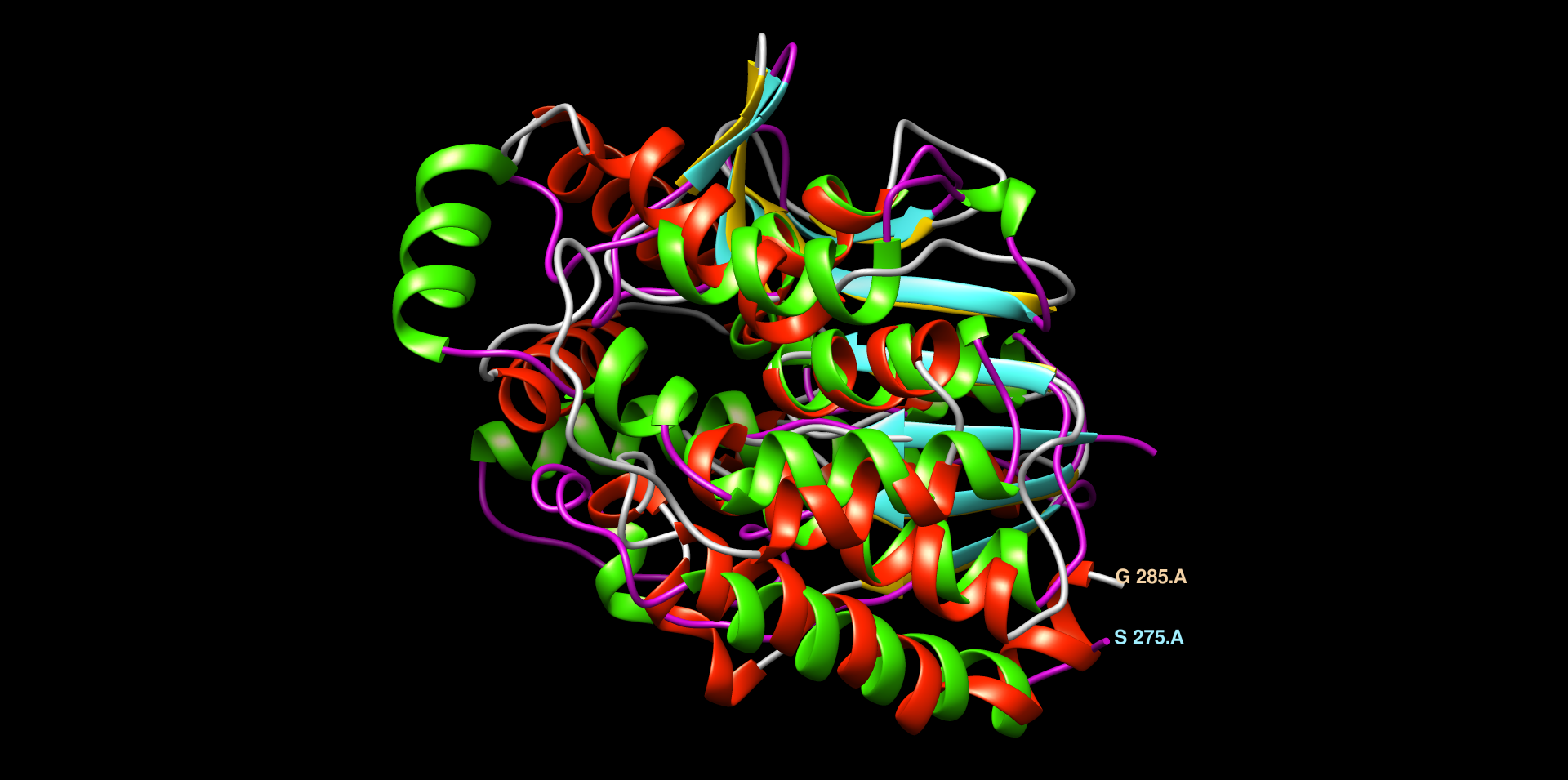
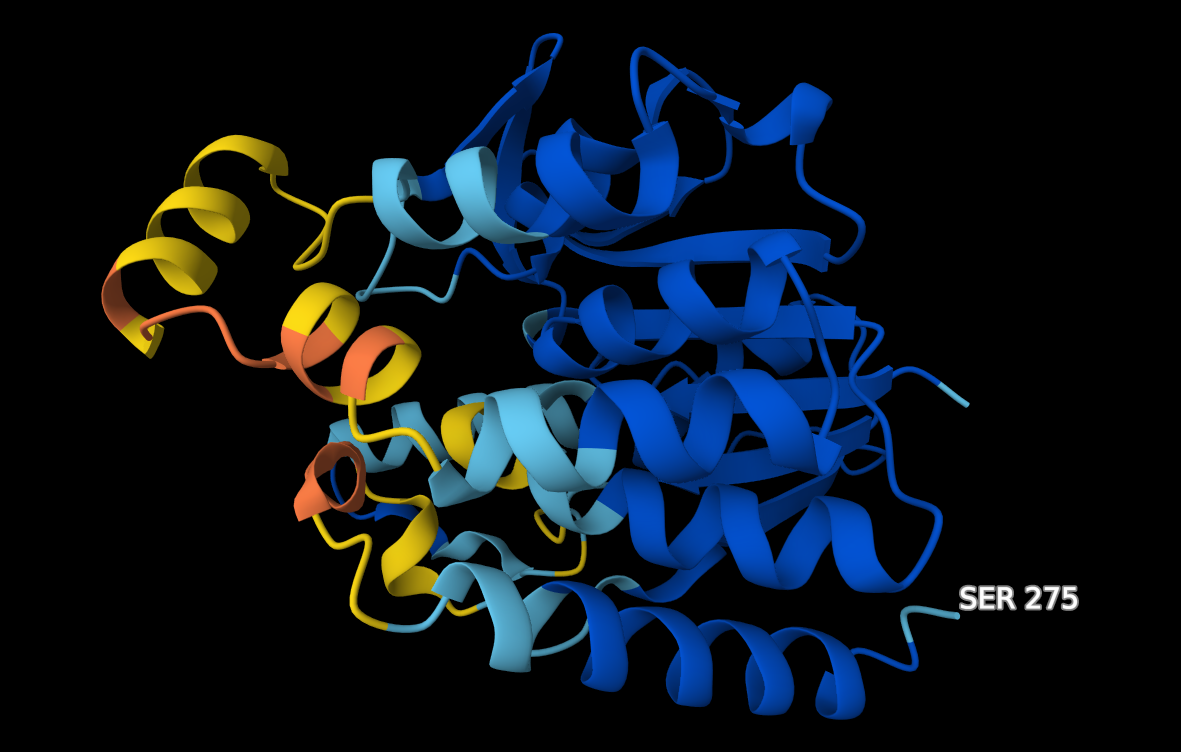
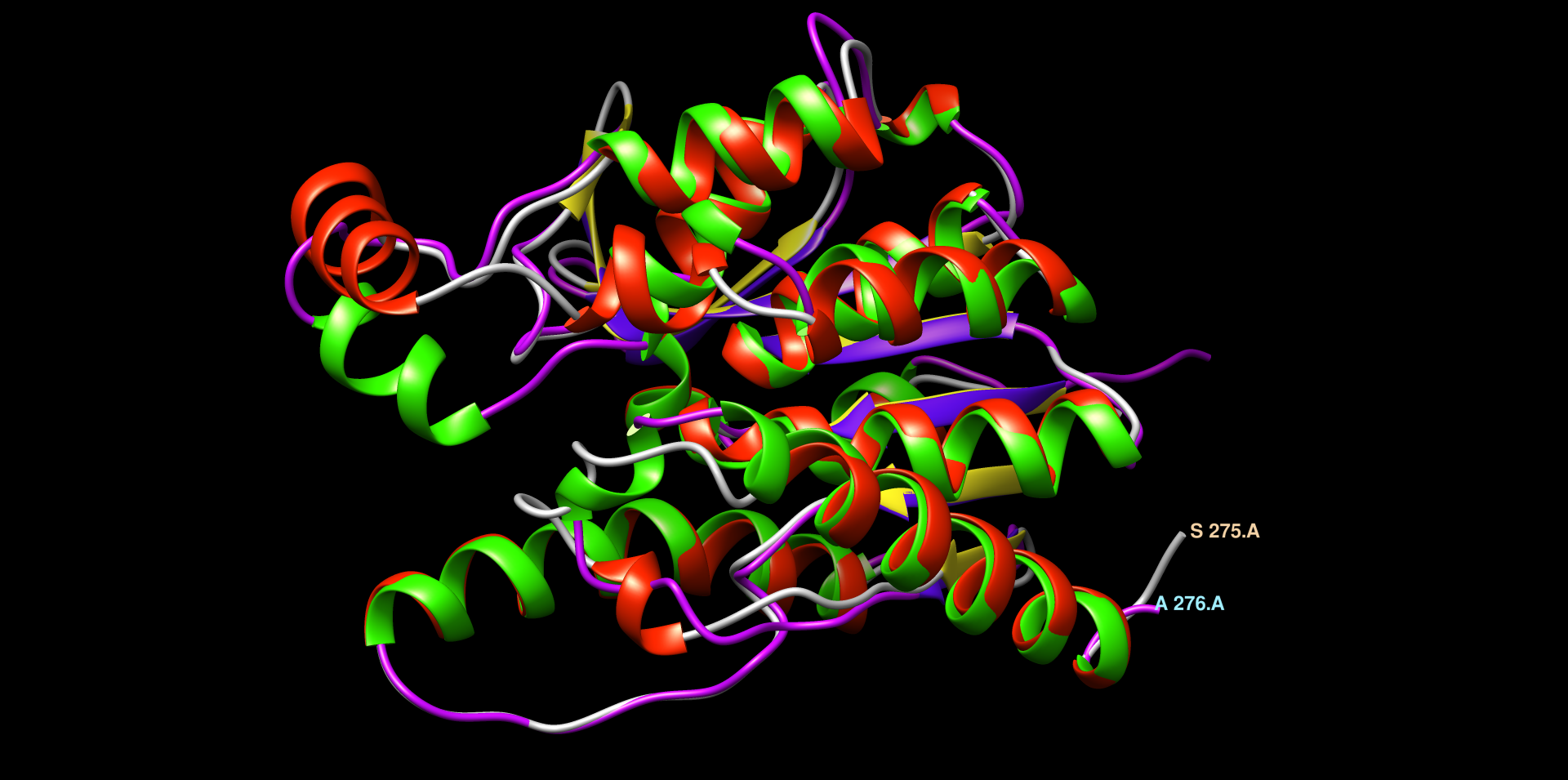
V chimeri sem uredil model strukture za YTR9. Moder, veliko alfa-vijačnic in zank, ne vidi se dobro beta-ploskev ! struktura. Naredil sem globalno poravnavo modelov struktur patatinu podobnih proteinov iz bakterij dveh različnih rodov ! poravnava_struktur. Pri tej poravnavi so dobro vidne tudi beta-trakovi. Strukturi sta dobro poravnani (LVT1 in YTR9). V MolStar sem tudi prenesel datoteko iz AlphaFold, da sem prikazal obarvanost modela strukture proteina YTR9 glede na zanesljivost ! struktura. Vidno je, da je struktura dobro ohranjena, večina elementov sekundarne strukture je temno modro obarvanih, kar nakazuje na visoko stopnjo zanesljivosti. Poravnav sem tudi strukturi YTR9 in M2M9, ki je podobna poravnava kot prejsnja, strukturi sta zelo dobro poravnani, torej ohranjeni ! poravnava_struktur.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
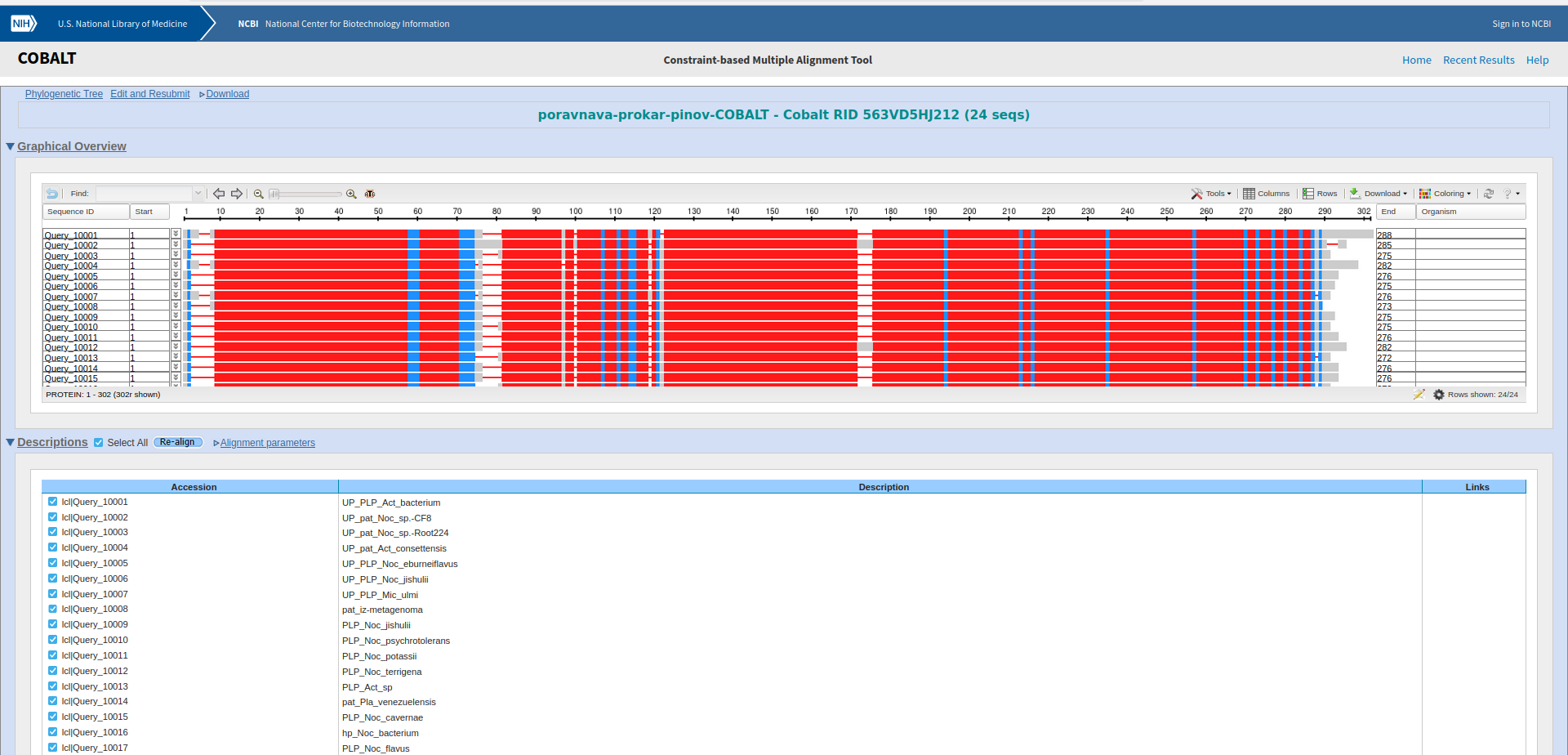
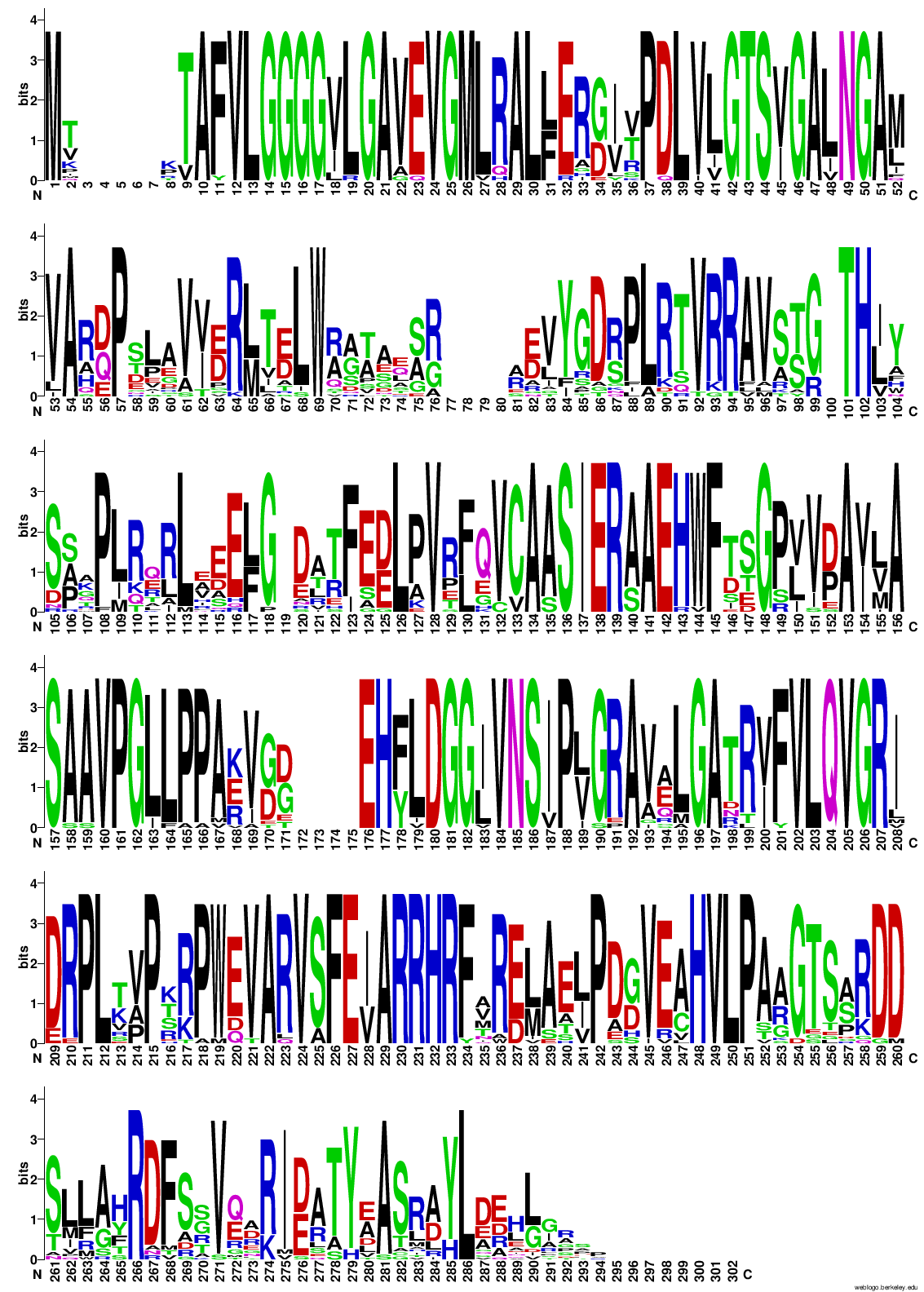
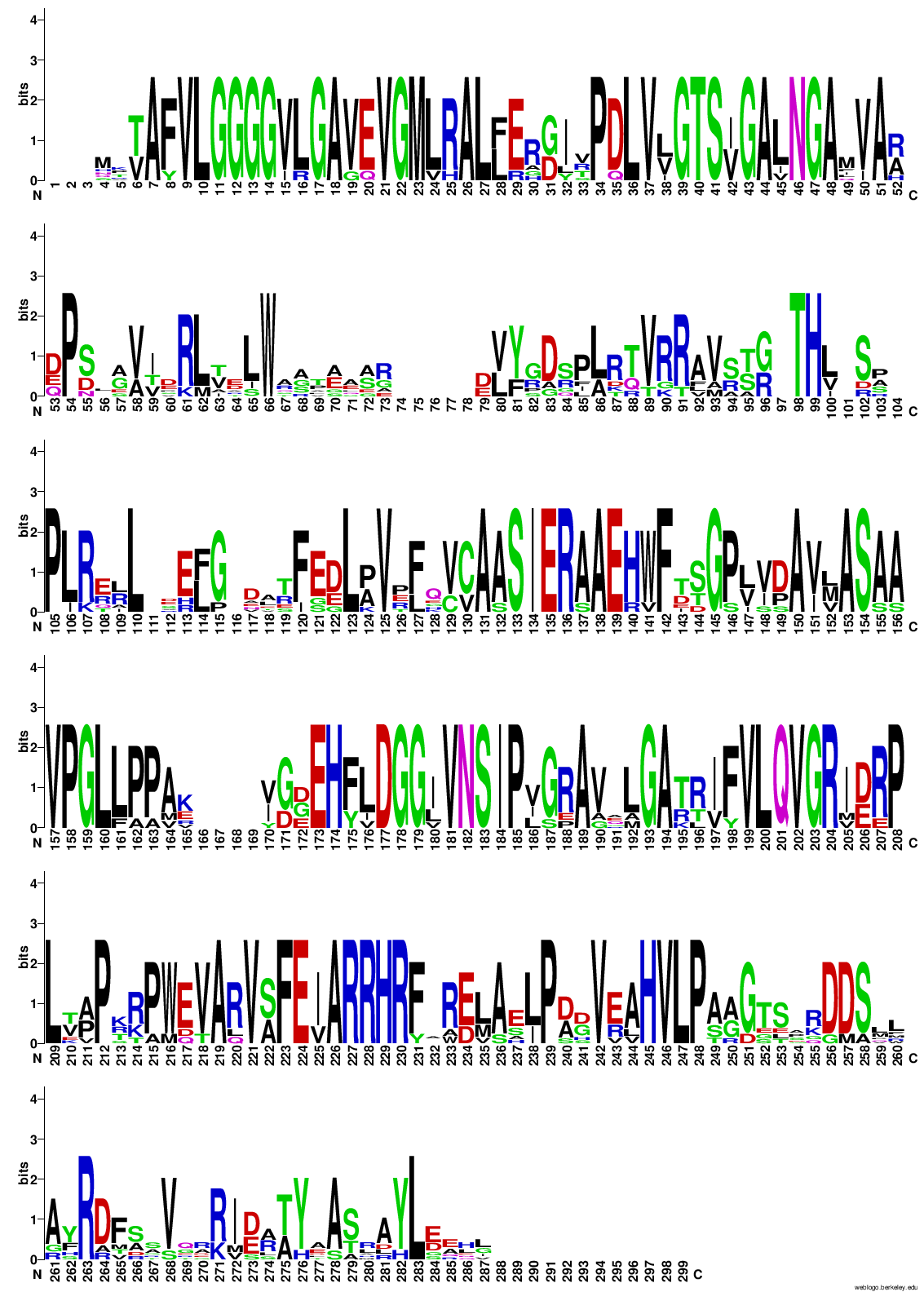
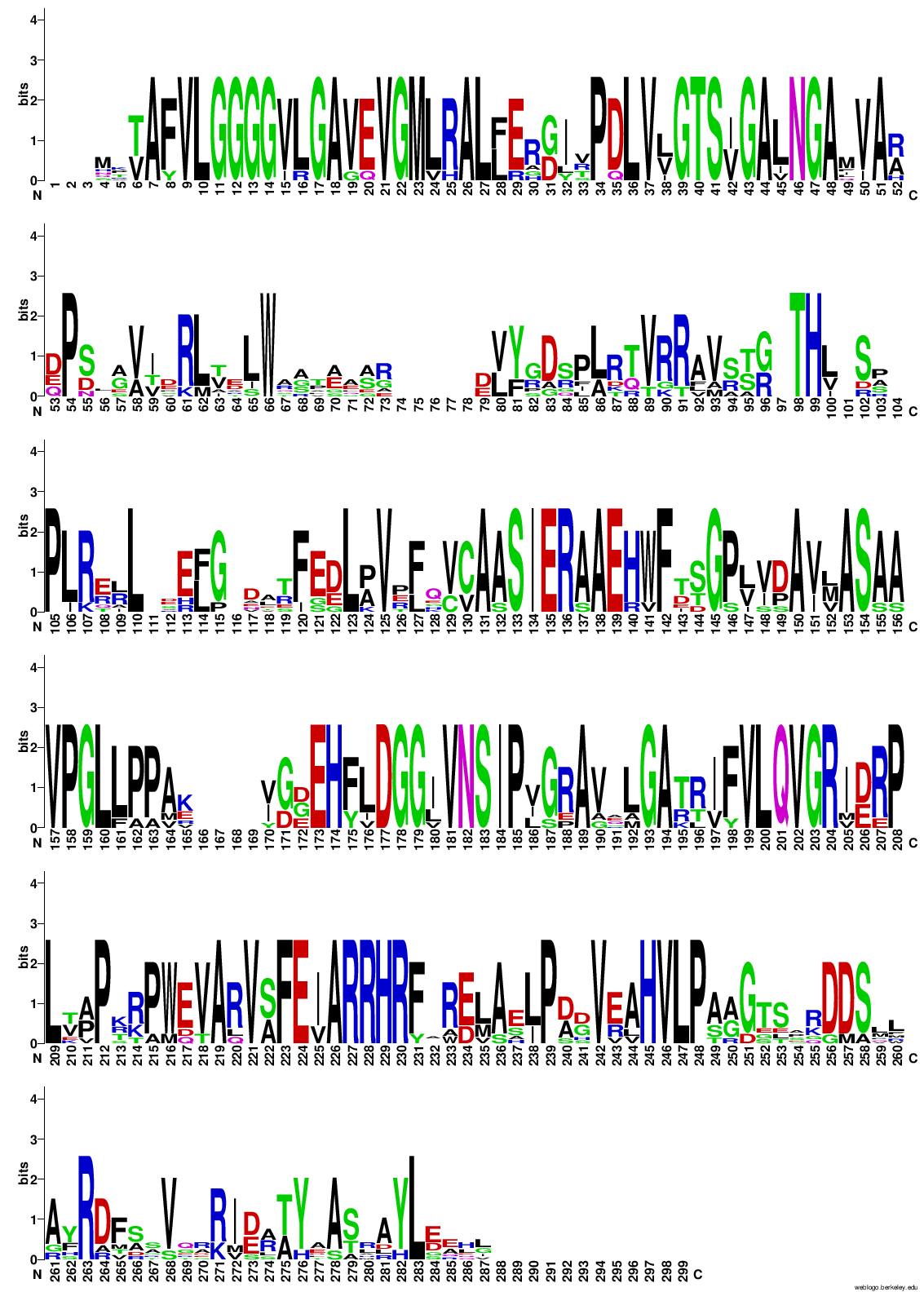
Potem sem z vsemi zaporedji, ki sem jih uporabil za izris drevesa, naredil poravnavo v orodju COBALT ! poravnava in podatke poravnave vnesel v WebLogo, da bi pogledal ali so kateri aminokislinski ostanki bolj ohranjeni, in bi morda lahko sklepal, da taki sodelujejo pri encimskem mehanizmu. A iz sequence logo-ta nisem mogel sklepati na aminokislinske ostanke, ki bi lahko bili pomembni za katalizo, ker je res veliko aminokislinkih ostankov dobro ohranjenih po skoraj celotnem zaporedju proteinov ! sequence_logo. Zgolj na začetku, na koncu in pri dveh regijah vmes v zaporedju proteinov ni dobro ohranjenih aminokislinskih ostankov.
{kind=link}
{kind=link}
Naredil sem tudi poravnavo v orodju COBALT-u samo z zaporeji proteinov, ki so podobni patatinu, ki sem jih našel na UniProt-u ! poravnava, nato pa sem podatke poravnave ustavil v orodje WebLogo za izris sequence logota. Zanimivo je bilo, da je izgledalo, kakor da je precej aminokislinkih ostankov v manjši meri ohranjenih, a je še zmeraj bilo precej načeloma dobro ohranjenih aminokislinskih ostankov ! sequence_logo.
{kind=link}
{kind=link}
Na UniProt-u sem tudi poravnal štiri proteine iz bakterij precej podobne patatinu, da sem prikazal kje se bi lahko nahajala transmembranska regija, če bi bila prisotna tudi pri ostalih proteinih ! poravnava. V UniProt-u sem našel zapis za to zgolj pri enem proteinu.
{kind=link}
Primerjava z evkariontskimi proteini#
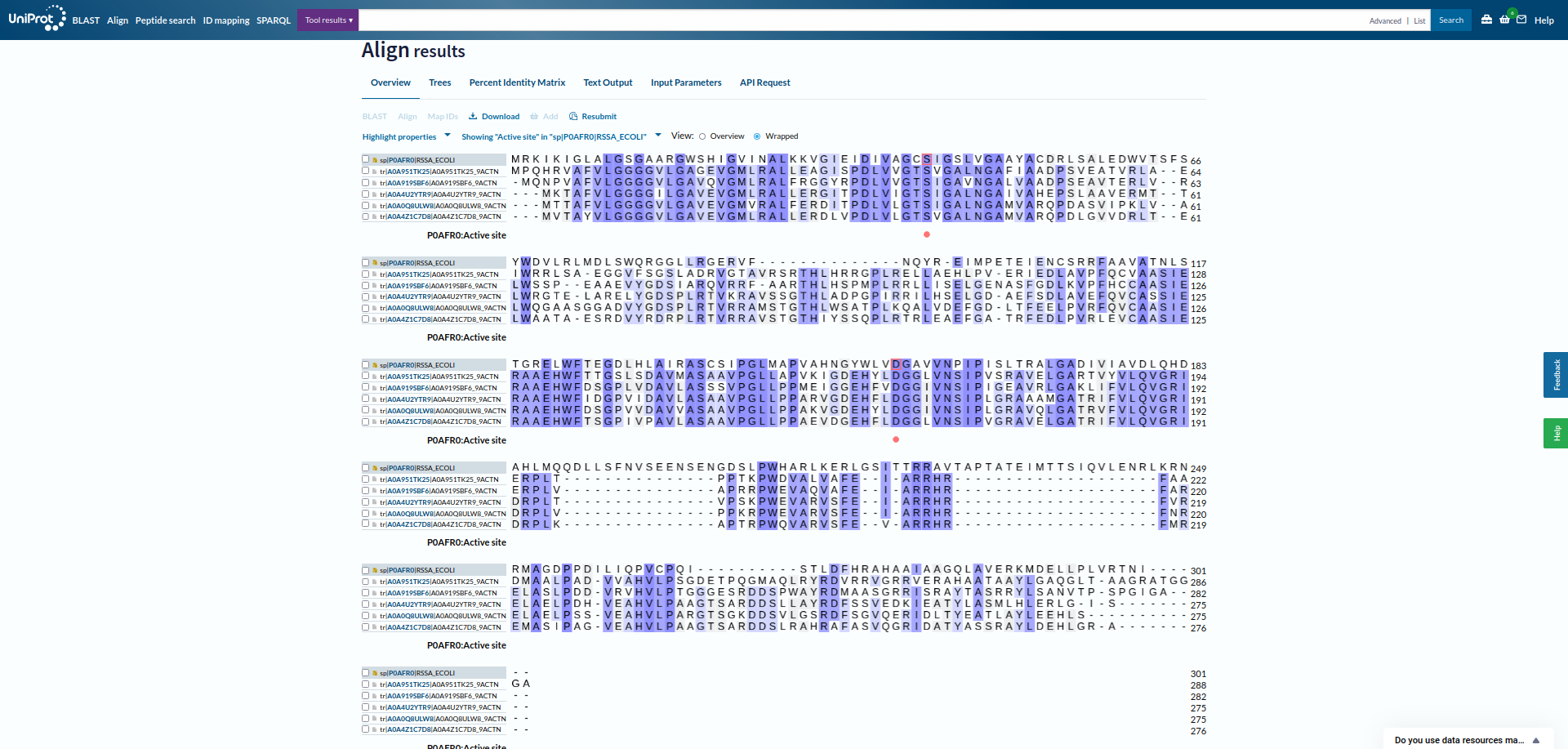
V blastp sem iskal z zaporedjem patatina anotirana zaporedja ! blastp. Našel je večinoma evkariontske proteine, vendar je med njimi tudi bil protein iz E. coli z domeno PNPLA, ki je bila pozicionirana približno na enakem mestu kot pri patatinu zelo podobnih proteinih. Skratka, tudi protein iz E. coli je bil precej podoben patatinu. Prejšnje, iz bakterij, zelo podobne proteine, sem poravnal skupaj z anotiranim proteinom iz E. coli v UniProt, da sem lahko označil aktivno mesto, in iz poravnave se zdi, da so tudi pri ostalih proteinih v aktivnem mestu lahko enaki aminokislinski ostanki ! poravnava. Protein iz E. coli je encim hidrolaza, in je udeležena pri razgradnji lipidov. Zelo podobne bakterijske proteine in protein iz E. coli sem poravnal tudi v orodju COBALT ! poravnava in naredil sequence logo z orodjem WebLogo ! sequence_logo. Ta se načeloma ni razlikoval od prejšnjega. Protein YTR9 (rdece barve heliksi, rumene ploskve in sive zanke) in protein iz E. coli (zeleni heliksi, modre ploskve in vijola zanke) sem s strukturami globalno poravnal v Chimeri. Razvidna je dobra ohranjenost strukture, saj se strukturi dobro pokrivata ! poravnava_struktur.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Vsi proteini podobni patatinu iz evkariontskih organizmov vsebujejo domeno PNPLA, ki je locirana bolj proti koncu zaporedja proteina. Vsi ti proteini imajo encimsko aktivnost, sposobni so hidrolizirati vezi. Prav tako vsi imajo vlogo pri metabolizmu lipidov, natančneje, razgradnji lipidov. V aktivnem mestu sta pri vseh proteinih prisotna S in D, ki sta pri vseh locirana na domeni PNPLA.
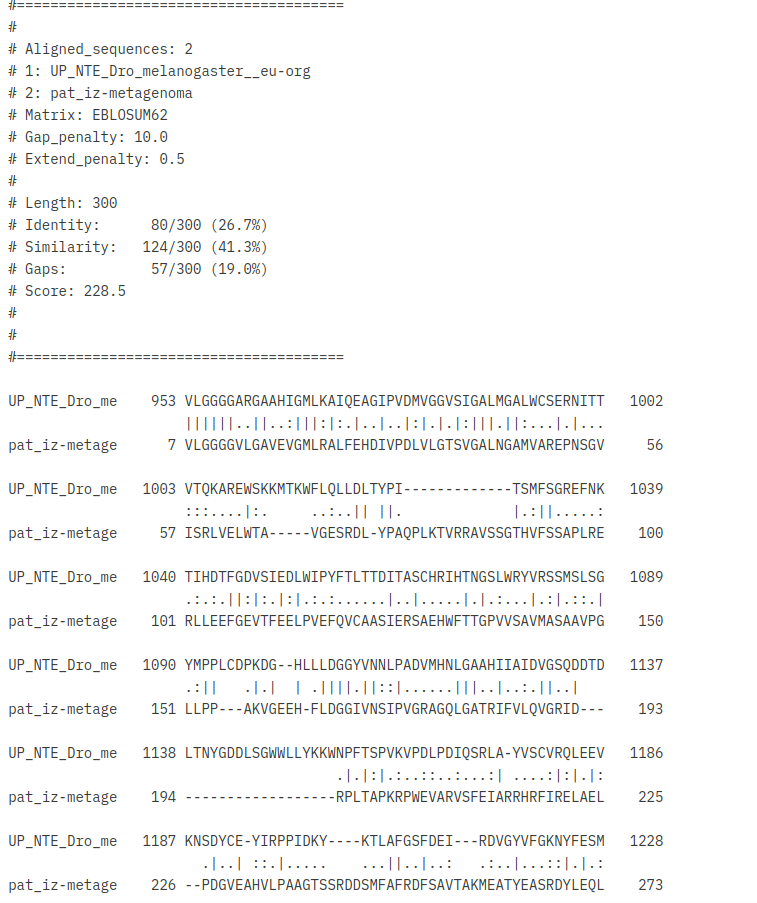
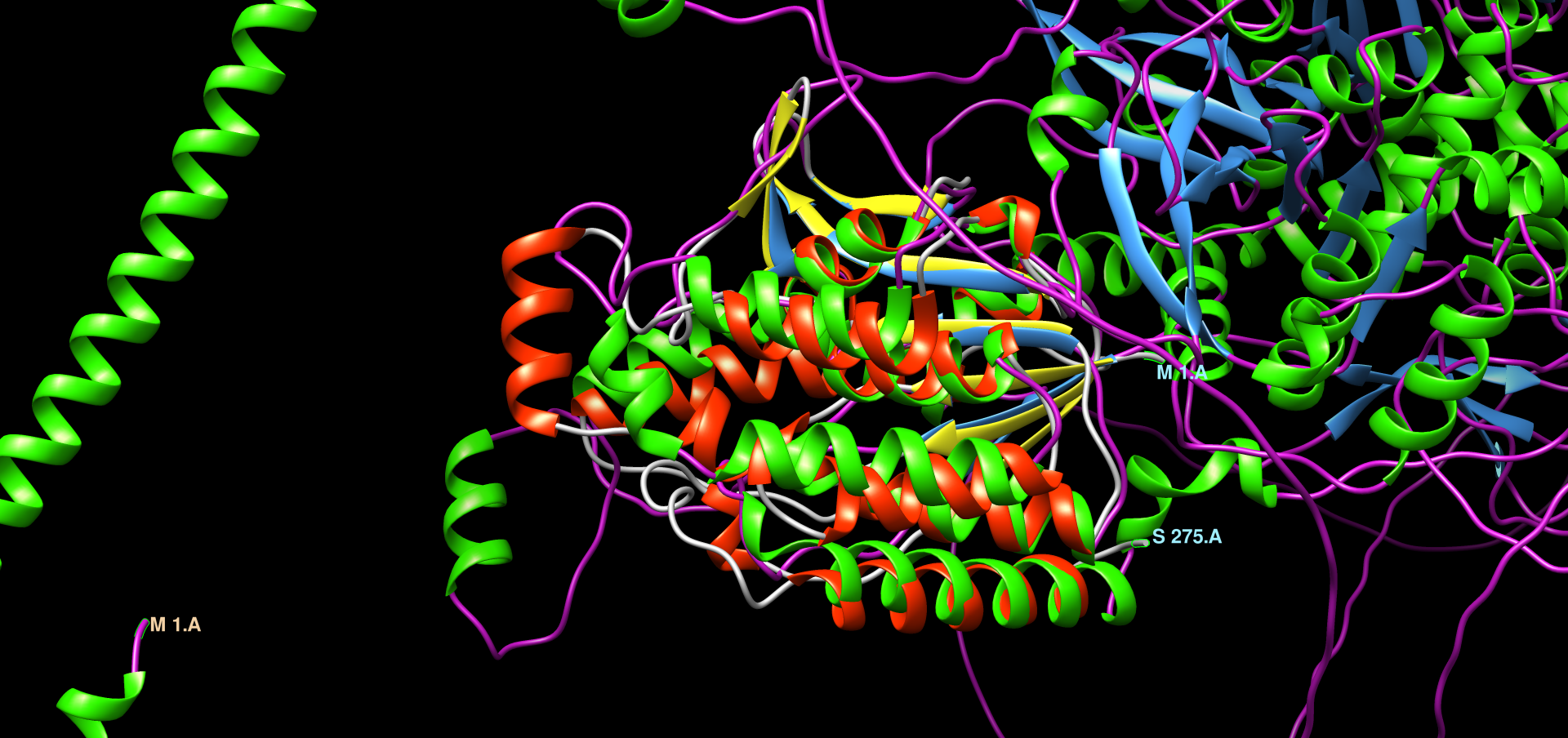
Patatinu najbolj podobni evkariontskimi proteini so veliko daljši, torej imajo veliko daljše aminokislinsko zaporedje od patatina, vendar se očitno patatin z evkariontskimi proteini dobro ujema v določeni regiji. Želel sem ugotoviti, kako dobra je poravnava struktur in zaporedij evkariontskih proteinov s proteinom patatinom. Zato sem si za primerjavo izbral patatinu podoben protein iz organizma Drosophila Melanogaster (vinska mušica). V orodju embossWater, ki uporablja določen algoritem za lokalno poravnavo zaporedij, sem lokalno poravnal zaporedje proteina patatina z zaporedjem proteina iz vinske mušice ! poravnava. Zaporedji se tudi lokalno ne ujemata tako zelo dobro, vendar bi rekel, da tudi ne tako slabo. Ujemata se v regiji, kjer je pri vinski mušici prisotna domena PNPLA, zato lahko sklepam, da je ta v določeni meri ohranjena tudi pri evkariontskih organizmih. Poravnavo struktur omenjenih dveh proteinov sem naredil v programu Chimera. In sicer je to bila lokalna poravnava z algoritmom embossWater. Strukturi se lokalno dobro prekrivata ! poravnava_struktur ! poravnava_struktur.
{kind=link}
{kind=link}
{kind=link}
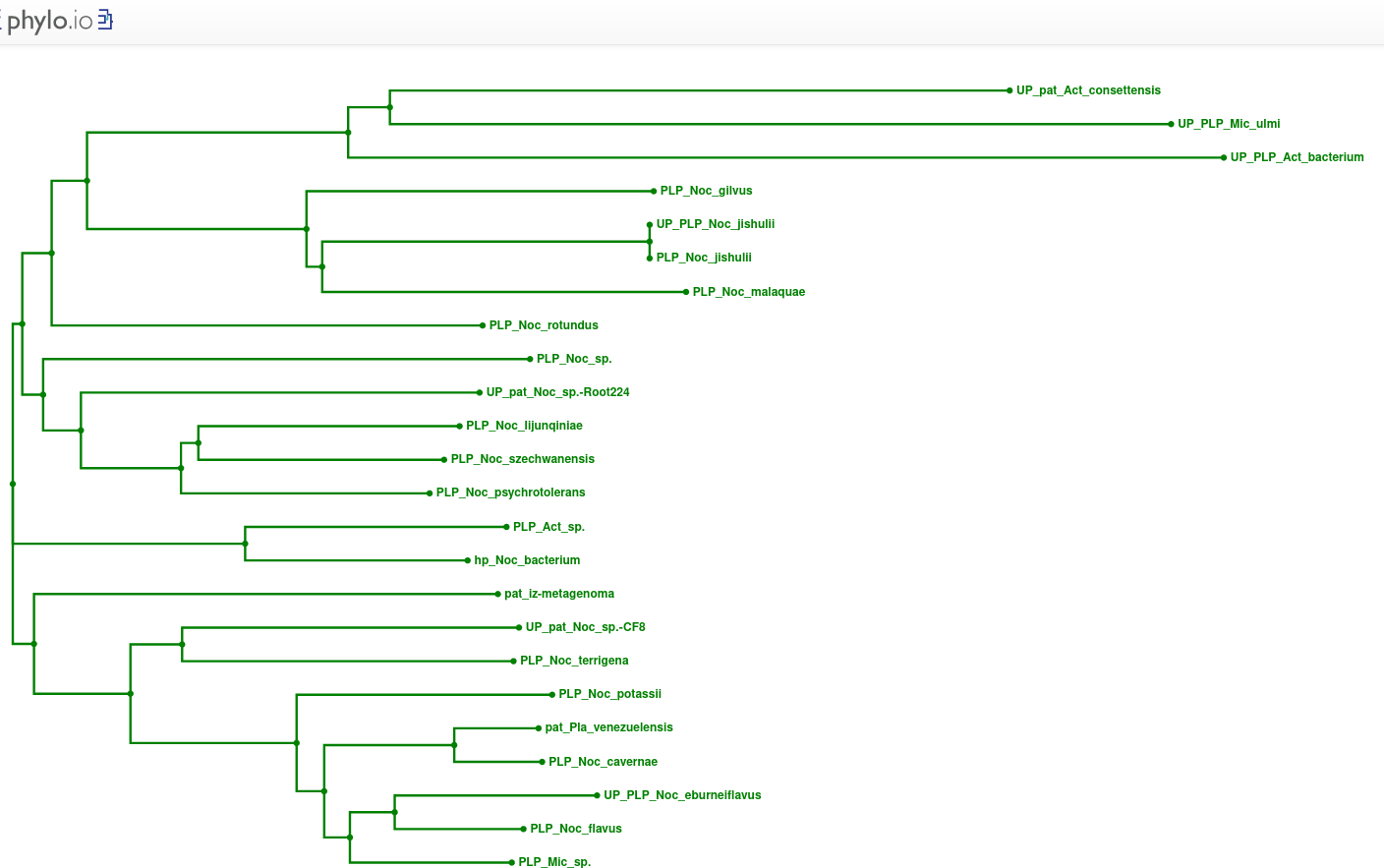
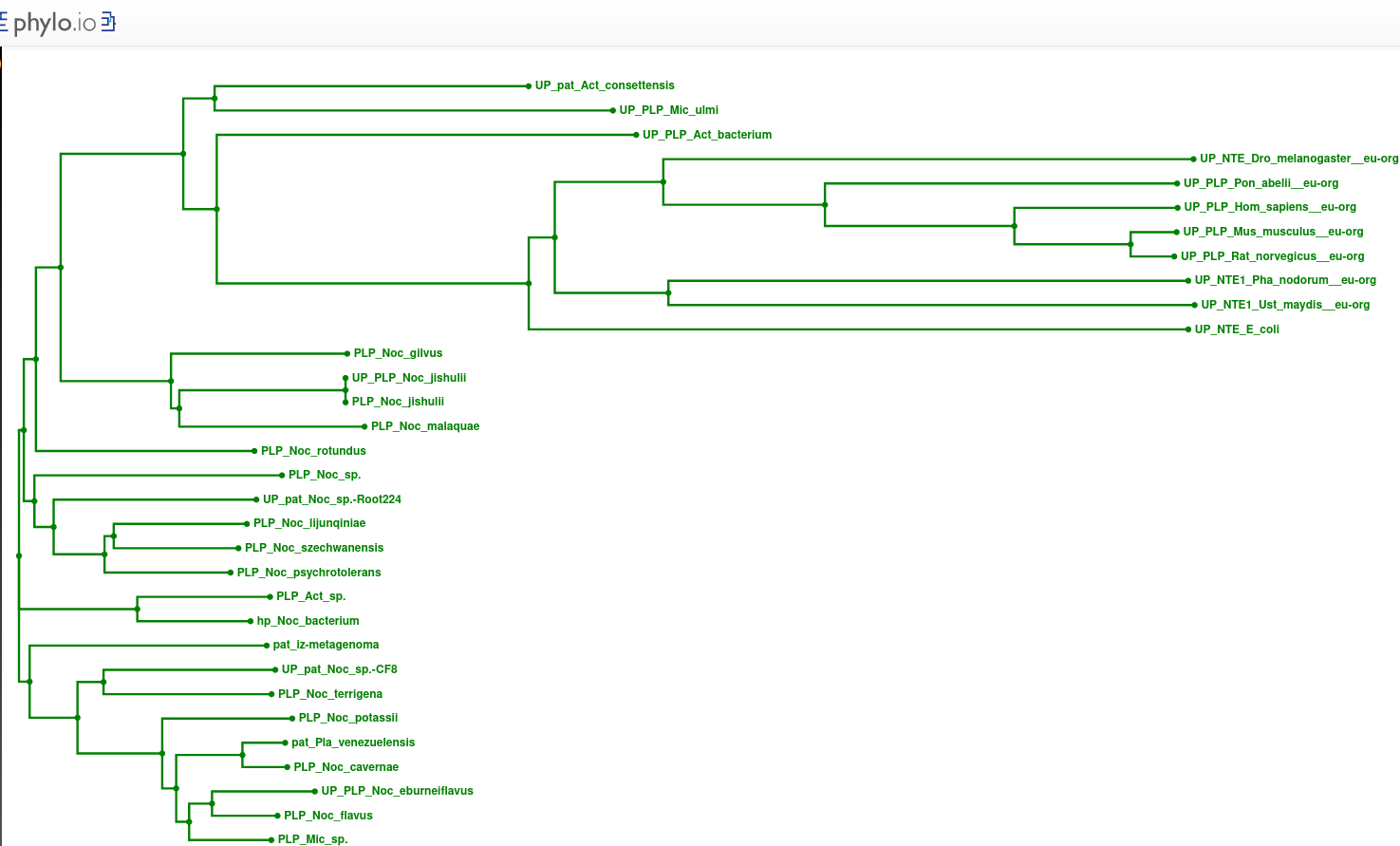
Zaporedja proteinov patatinu podobnih proteinov iz bakterij in iz evkariontskih organizmov sem poravnal z orodjem Clustal Omega ! poravnava, potem pa sem s podatki poravnave, uporabne za izris drevesa, narisal filogenetsko drevo v orodju phylo.io ! filogenetsko_drevo.
{kind=link}
{kind=link}
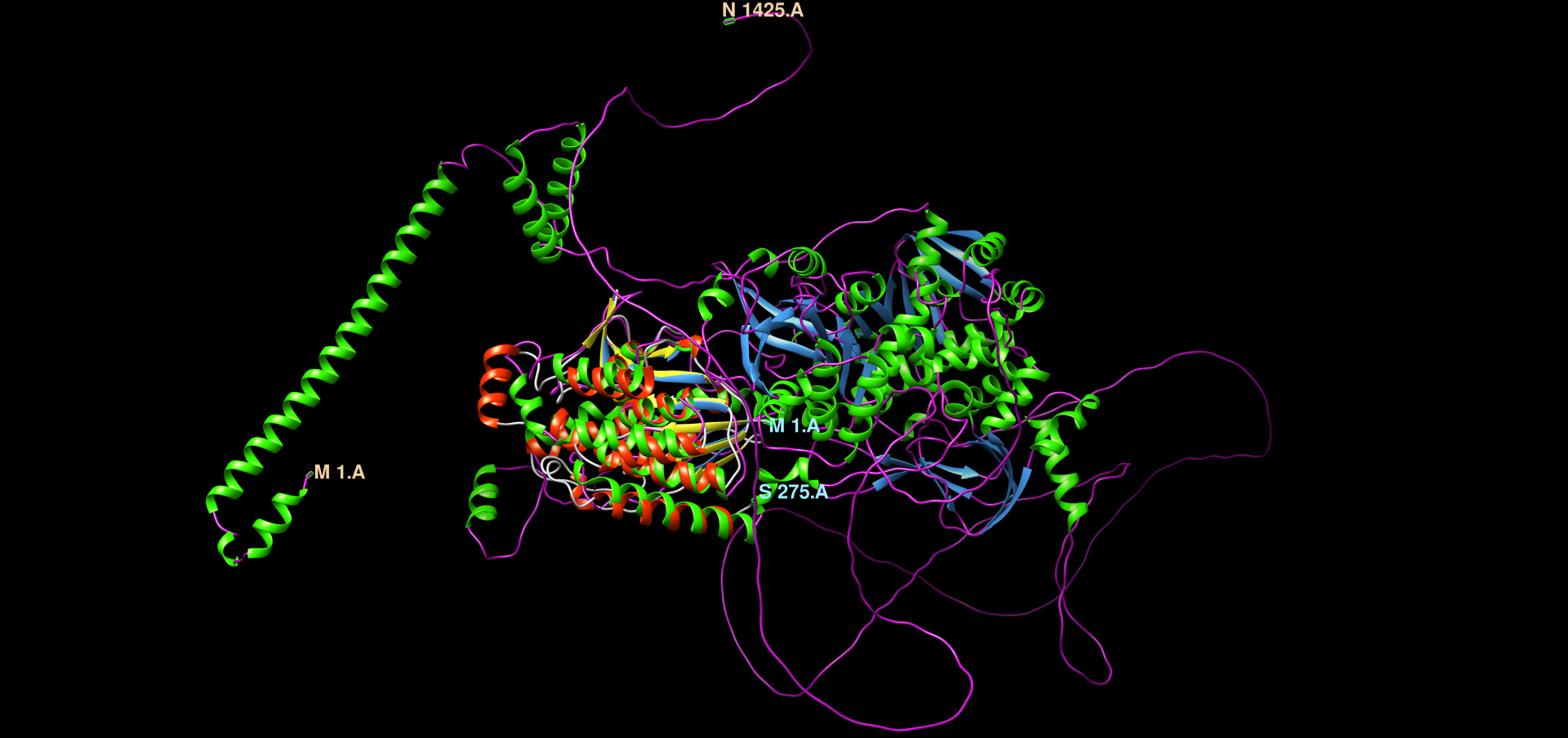
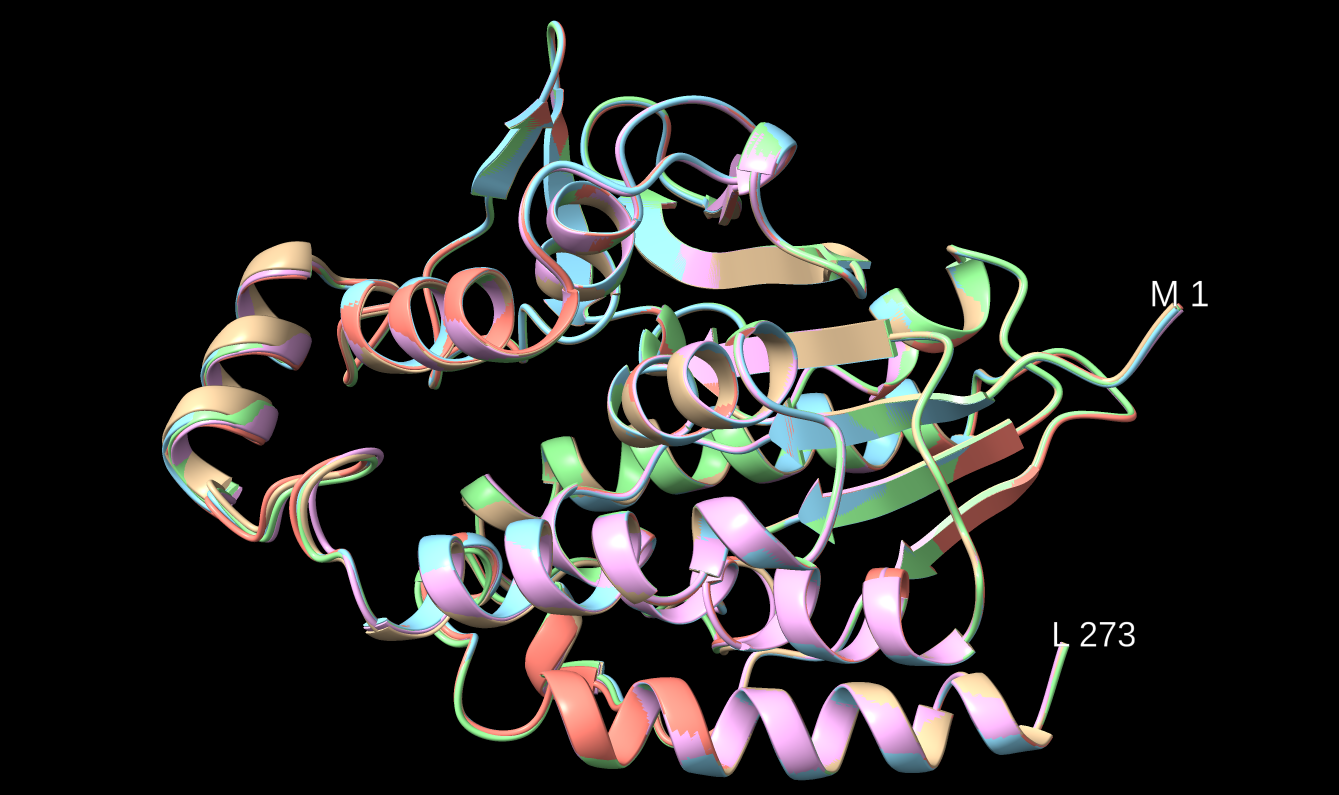
Na AplhaFold Server sem ustavil zaporedje patatina iz metagenoma za napoved strukture. Ko je program izrisal model strukture, sem naložil datoteke, v katerih je bilo pet različnih modelov struktur. V ChimeriX sem naredil superpozicijo teh petih modelov struktur ! superpozicija_struktur. Strukture so precej prekrite med seboj, dobro so ohranjene. Iz alphafold serverja sem slikal model pobarvan po zanesljivosti strukture, ki tudi prikazuje podobno ohranjenost strukture, kot sem jo izrisal z MolStar za patatinu podoben protein ! struktura.
{kind=link}
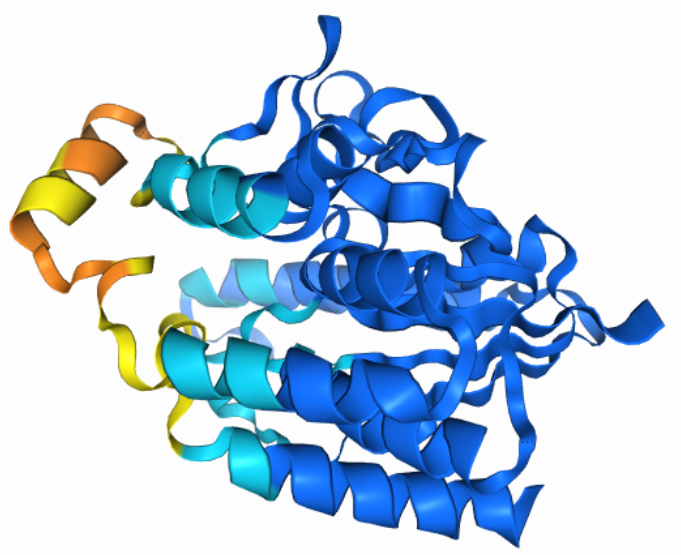{kind=link}
Na koncu sem naredil tudi poravnavo in prikaz aktivnega mesta na proteinu iz E. coli in možnega aktivnega mesta na patatinu ! poravnava_struktur ! poravnava_struktur.
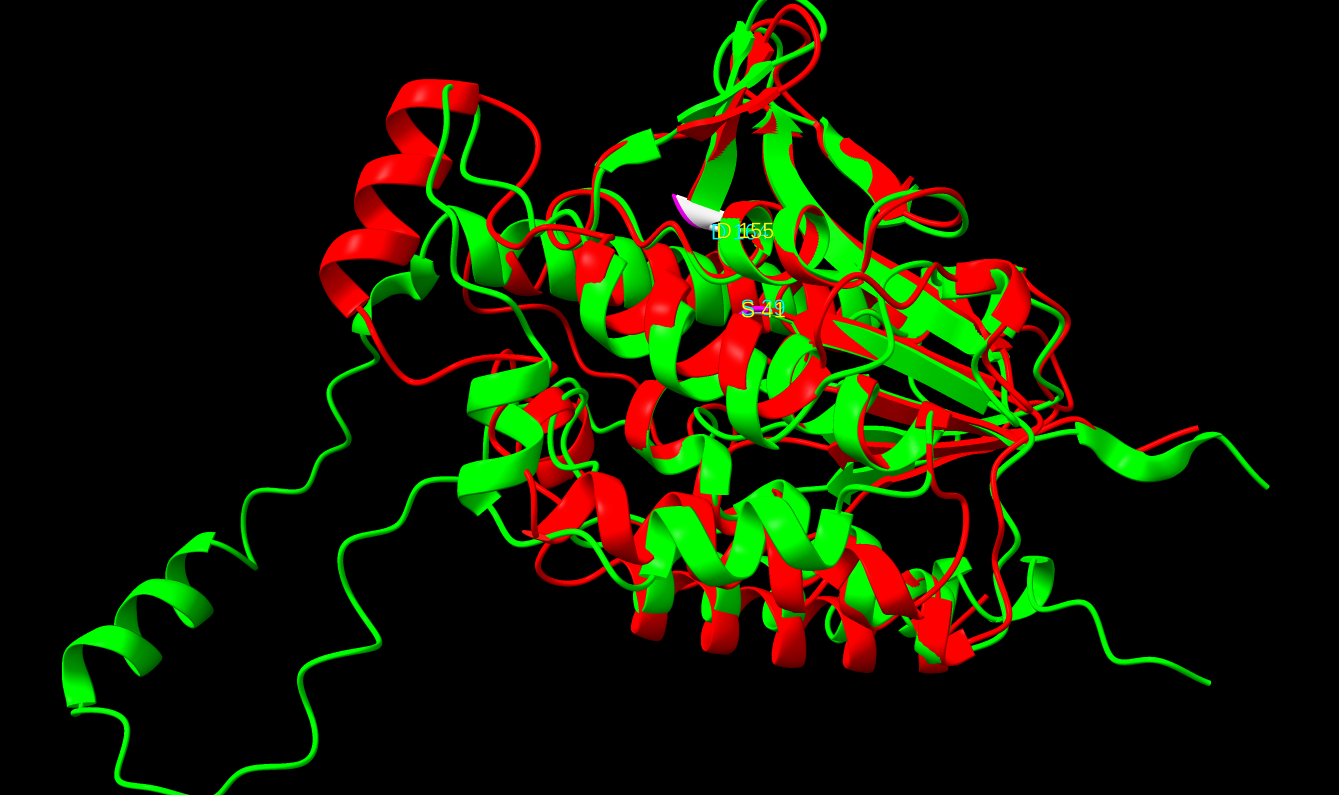{kind=link}
{kind=link}